この講義の内容は下記のYouTubeサイト(生命科学を専攻する学生の為の分子生物学講義)で視聴することができます
https://www.youtube.com/watch?v=p_jyIe3gqh4
1. この講義の内容について
この動画では,リボソームを構成するrRNAやリボソームタンパクについて解説しています。
- 大サブユニット(Large subunit),小サブユニット(Small subunit)を構成するrRNAやタンパクの配置について説明しています。
- またrRNA遺伝子をコードするオペロン(operon)やリボソームタンパクをコードするオペロンについても言及しています。
- ペプチド鎖の伸長反応におけるリボソームの大小サブユニットの動きとペプチジルtRNA, アミノアシルtRNAのリボソーム内での位置についても詳しく説明しています。
- また,rRNAがペプチドの転移反応を触媒する活性を持つリボザイム(ribozyme)である事を示し,そのサイトを23SrRNA中に示しています。
Key Words: E-site,P-site, A-site, Large subunit, Small subunit,5 S rRNA, 16S rRNA, 23S rRNA, Ribozyme, Peptidyl-tRNA, Aminoacyl-tRNA, Large Subunit, Small subunit, Aminoacyl-tRNA, Peptide transfer, Nucleolus, Ribosome assembly, L7/L1235S rRNA, Ribosomal RNA operon, tRNA, hybrid site
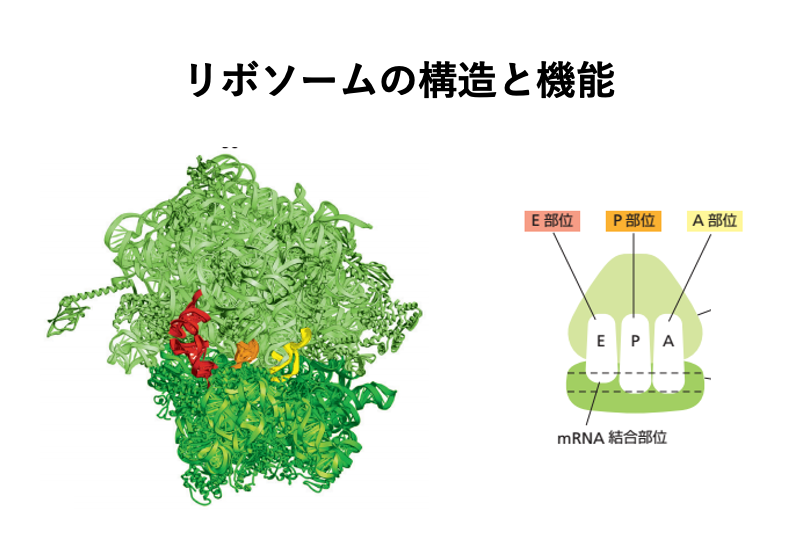
2. リボソームの基本構造
リボソームは、コドンによって指定された順番でアミノ酸がつなぎ合わされることで、 遺伝子の情報にったペプチド鎖を生産する装置ということができます。
この反応を スムースに実施するために,リボソームには,E-site, P-site, A-siteと呼ばれる3つのtransfer RNAを格納できる空間があます。
具体的には、アミノ酸を付着させたtransfer RNAが、 そのアンチコドンによりコドンと安定な水素結合を作りながら、リボソームの中でペプチド鎖を伸長させることで、それが実現されています。
リボソームには、ラージサブユニットと呼ばれる大きな部分と、スモールサブユニットと呼ばれる2つの部分があります。 こラージサブユニットとスモールサブユニットは、メッセンジャーRNAを,2つのサブユニットの境界面に配置して,1つの複合体を形成しています。
3. リボソームの基本構造:真核生物と原核生物での比較
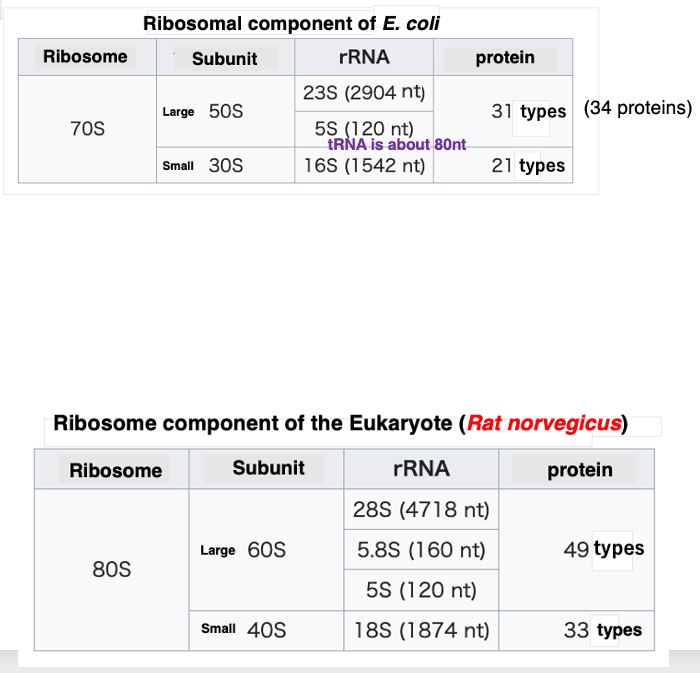
最初に、細菌である大腸菌のリボソームがどのように構成されているかを述べます。 大サブユニットは50SというSedimentation coefficientを持っていて、小サブユニットは30Sと呼ばれるSedimentation coefficientを持っています。 このSedimentation coefficientが大きければ大きいほど、その粒子は大きな分子であって早く沈むということを示しています。このように粒子の大きさがSedimentation coefficientを使って表現されるのは、リボソームの研究の初期において、遠心機を使い,沈降速度の差を利用して粒子が分けるという実験手法が頻繁に用いられていたという歴史的な背景があります。Large subunitはRNAとタンパクの複合体であるということがわかっています。 この粒子内には2種類のRNAが内蔵されています。大きい方のRNAのSedimentation coefficientは23Sであり, 2904塩基から構成されています。小さい方のSedimentation coefficient は5Sであり,120塩基からなります。 また、31種類34個のタンパクが,この粒子には含まれています。つまり,31種類34個のタンパクと2904ヌクレオチドからなる23SリボソームRNA、120塩基からなる5S RNAの複合体がLarge subunitということになります。
一方、30SというSedimentation coefficientを持つSmall subunitは、16Sと呼ばれる1542塩基からなるRNAと、21種類21個のタンパクの複合体です。50Sと30Sの粒子が会合すると、Sedimentation coefficient が70Sのリボソームとなります。 ここで,Sedimentation coefficientではなくて,リボソームの大きさを質量で表現しましょう。Large subunitは、1600 kilo Daltonという質量を持ちます。 30S Small subunitは、900 kilo Daltonという質量になることがわかります。 平均的なタンパクの大きさというのは、およそ300個のアミノ酸がつながったものですけれども、その質量はおよそ33 kilo Daltonになります。このことから,1600 kilo Dalton の質量を持つ70S Large subunitが如何に大きいものかというのがわかるかと思います。 Transfer RNAは80塩基弱で構成されているRNAで、Sedimentation coefficient としては4Sです。5S ribosome RNAはtransfer RNAよりは少し長い120塩基で構成されています。 それらと比べると、23Sや16S RNAは巨大なRNA分子だということがわります。
次に、真核細胞に含まれるリボソームについても見ていきたいと思います。 これはラットのリボソームに関する表です。 これも細菌と同じように、Large subunitとSmall subunitという2つのサブユニットで構成されています。 Large subunitのSedimentation coefficient は60Sであり、Small subunitのSedimentation coefficient は40Sなので,原核生物のサブユニットよりは,すこし大きくなっています。 Large subunitには,8S、5.8S, 5SのSedimentation coefficient を持つ3種類のリボソームRNAが含まれています。原核生物の場合には、Large subunitには2種のリボソームしか含まれないのに対して、 真核生物の場合では,5.8S RNAが追加されて3種類となっています。また、Large subunitに含まれるタンパクは49種、49個のタンパクです。 一方、Small subunitのSedimentation coefficient は40Sです。 ここに含まれるRNAは一種類のみで、18S RNAと呼ばれます。それは,1874塩基からなるRNAです。 その他に33種、33個のタンパクがこのSmall subunitには含まれています。 60S Large subunit の質量は2800 kilo Daltonです。 40S Small subunitの質量は,1400 kilo Daltonです。この2つのサブユニットが会合すると、Sedimentation coefficient が80Sのリボソーム粒子となります。原核生物の会合したリボソームが70Sであるのに対して,それよりも,すこし大きな粒子となっています。
4. リボソームの立体構造の外観
では、リボソームの外観を見ていきましょう。
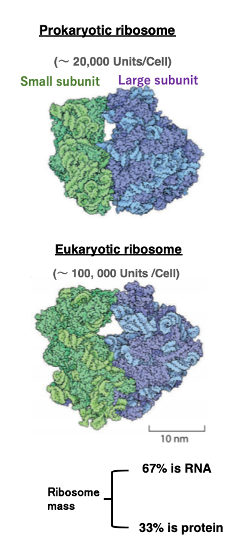
上図が細菌のリボソームの外観です。 このようなリボソームが、細菌ではおよそ細胞あたり2万個あります。
それに対して、下図が真核生物のリボソームの外観です。 真核生物では、リボソームが細胞あたりおよそ10万個程度あります。 緑色がsmall subunit、青色がLarge subunitです。
細菌と真核生物のリボソームを比べますと、 細菌のほうが構成タンパクの数も少ないし、 リボソームのRNAも少し短いく,全体の大きさも小さい感じがします。
リボソームを構成するのはタンパクとRNAですが,質量比で言いますと、 3分の2がRNA、3分の1がタンパクです。
5. 2D structure model of E. coli 16S rRNA
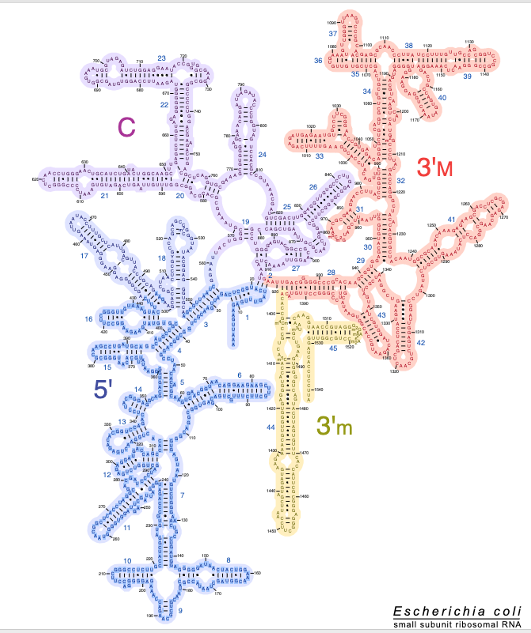
この図は大腸菌の16SリボソームRNAの二次元モデルです。
実際にはリボソームRNAは非常に複雑な三次元構造をとっています。 二次元構造ではヘアピンとして表現される,このような部分が 他のヘアピン部分と水素結合を形成することで 複雑に折りたたまれた三次元構造をとります。
6. 2D structure model of E. coli 23 and 5S rRNAs
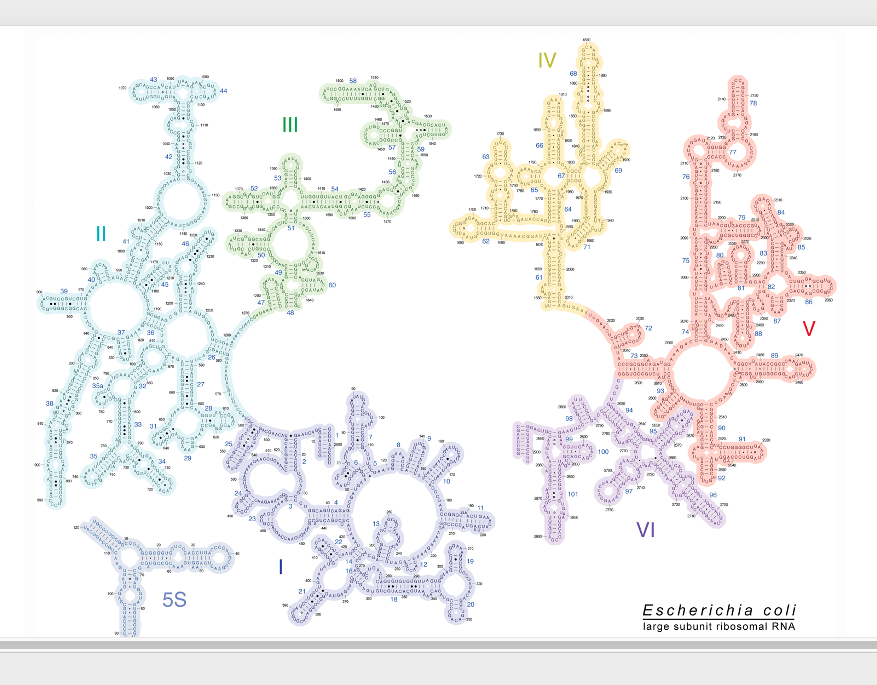
次に,これが大腸菌の23SリボソームRNAと5S リボソームRNAの二次元構造モデルです。
この図ではこことここが切れていますけど、実際にはつながっていますし、 この部分とこの部分が実際にはつながっている一本のRNA分子ということになっています。 これも実際には,非常に複雑に折りたたまれた三次元構造をとります。
ここに120塩基の短いリボソームRNAである5SリボソームRNAの二次元モデルが示されています。
7. Three-dimensional lstructure of a bacterial ribosome
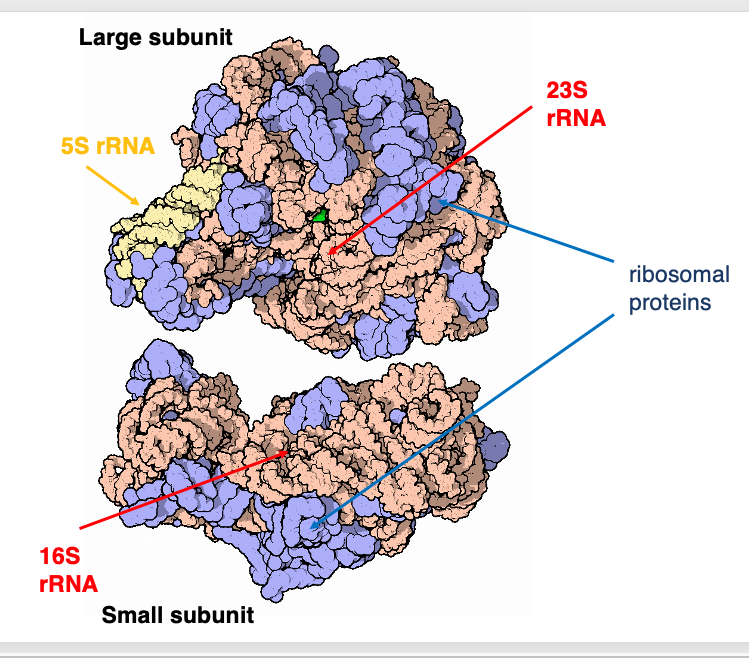
ではもう少し立体的なイメージで描かれたリボソムを見ていきたいと思います。 これは細菌のribosome RNAのLarge subunit。 こちらがSmall subunitです。 ここでは5S ribosome RNAは黄色で示されています。 また23S ribosome RNAはこのようにオレンジ色で示されています。 またribosome proteinsはブルーで示されています。
Ribosome proteinsは複雑な立体構造を持つribosome RNAの内部に配置されることはなく、 ribosome RNAの表面に配置されているということがわかっています。 こちらSmall subunitですが、16S ribosome RNAがピンクで示されています。一方で,ブルーはタンパクを示しています。 ラLarge subunitと同じようにSmall subunitでもribosome proteinsはribosome RNAが形成した複雑な立体構造の表面に配置されています。
ribosome proteinsの役割としては,ribosome RNAが正しい立体構造を形成したり,維持するのを助けたり, ribosome RNAが活性化するために構造を変化させるのを助けているという役割を担っていることが分かっています。リボソームが行っている様々な化学反応の活性中心はribosome RNAによって形成され,タンパクには活性中心が存在しないことが分かっています。
8. Three-dimensional image of E. coli ribosome with modified rRNA bases
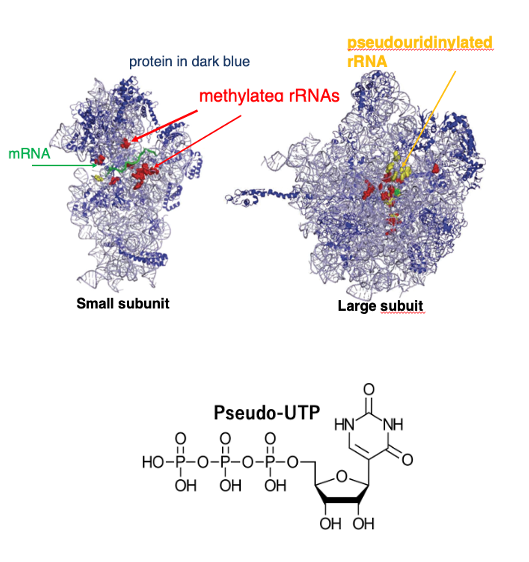
この図は、大腸菌のリボゾームの立体的なイメージを示しています。
左側がSmall subunit, 右側がLarge subunitを示しています。この図では、こういうふうな薄く描かれているのがリボゾームRNAです。 また、リボゾームタンパクは濃いブルーで描かれています。
リボゾームRNAでは、塩基がメチル化されている場合があります。 この図では、メチル化されたリボゾームRNA塩基の位置を赤で示しています。 また、塩基がシュードウリジン化されている場合があります。 そういうシ塩基は黄色で示しています。
RNAの塩基ばかりでなく、リボゾームタンパクも リン酸化やメチル化修飾のようは,翻訳後修飾を受けるということが知られています。このような一連の修飾は、リボゾームが正しく機能するために必須であるということも知られています。
9. Saccharomyces cerevisiaeの80Sリボゾーム
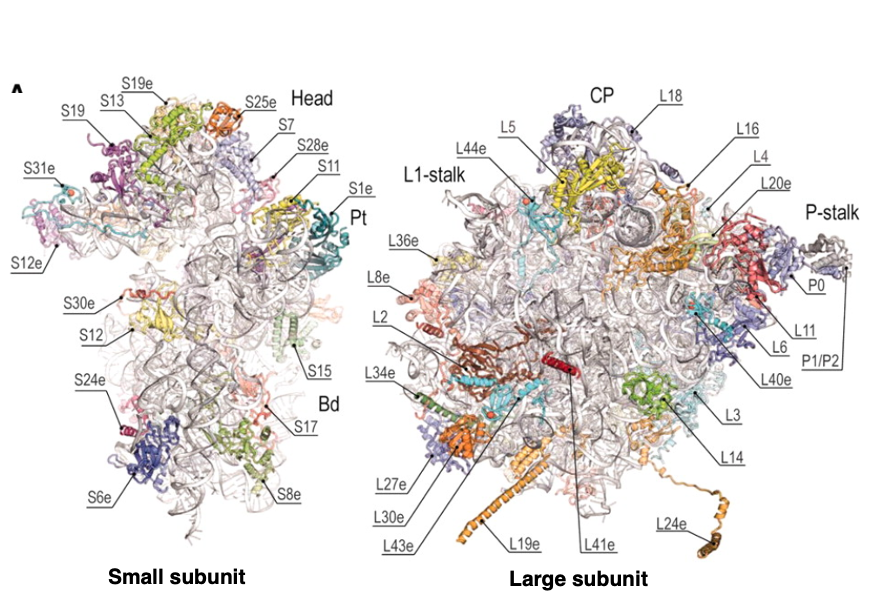
酵母の一種,Saccharomyces cerevisiaeの80Sリボゾームの立体構造図です。 左側がsmall subunit、右側がlarge subunitです。
この図では特にリボゾームRNAの表面に配置されているタンパクについてその位置が明示されています。 タンパクはRNAの表面についているということがよくわかると思います
10. E. coli ribosomal RNAオペロンの転写と加工
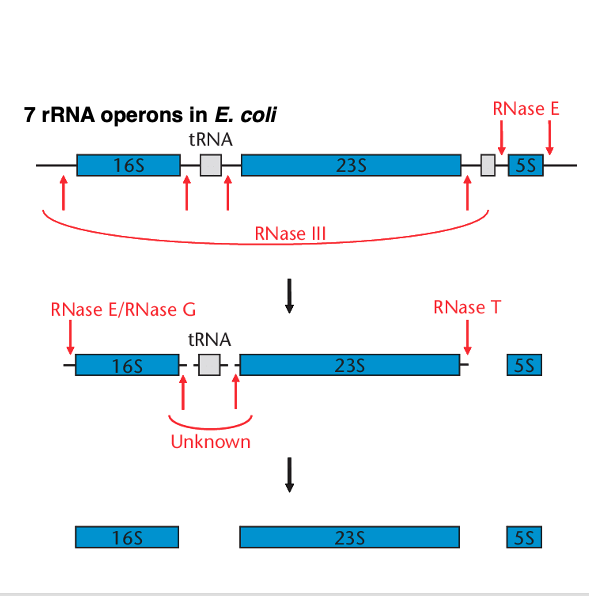
ここからは,リボソームの構成要素であるribosomal RNAや リボソームタンパクがどのようにして作られるのかについて解説をしていきます。
まずは,細菌のribosomal RNAオペロンの転写について解説をしていきます。 大腸菌では, リボソームRNAオペロンが7つありますが,どれも同様の構造をしています。
オペロンから転写されて作られた 長いRNAはRNase E, RNase G、RNase T などにより,分断されていきます。 最終的には16S ribosome RNA、23S ribosome RNA、5S ribosome RNA、transfer RNAに切断され,加工されます。
11. Ribosomal protein operons in E. coli
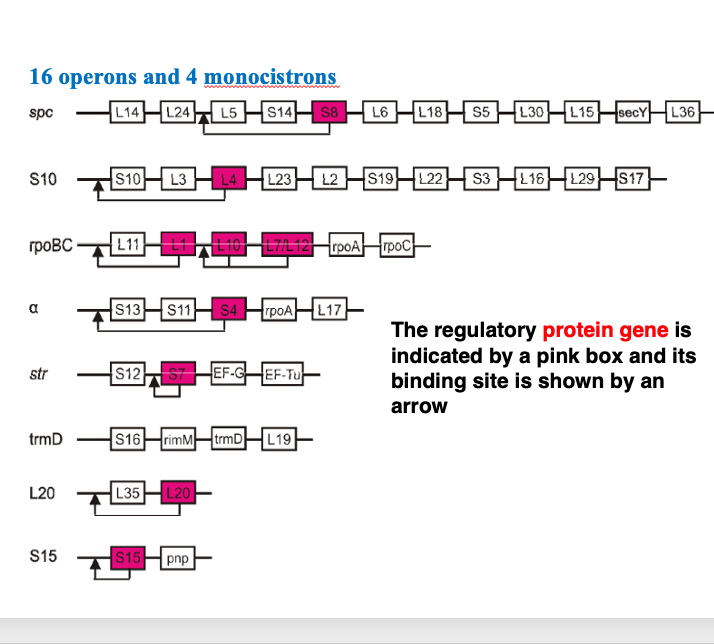
大腸菌のリボソームタンパクをコードしているオペロンについて解説をしていきます。 ここには代表的な8つのオペロンを載せています。
Spc-operonにあるS8リボソームタンパクをコードする遺伝子が赤色で示されています。このS8遺伝子の産物であるS8タンパクは、メッセンジャーRNAのこの部分に付着することによって、 負のフィードバックをかける制御タンパクとしても機能することが知られています。この図で示したように,リボソームタンパクをコードしているオペロンの転写制御は,その産物が自己をコードしているmessenger RNAの転写を制御するというcis-型のフィードバック制御が行われています。
リボゾムでは、それぞれのタンパクが基本的には粒子内に1つずつ配置されています。 そのために、それぞれのタンパクの量は、厳密に同個数ずつ生産される必要があります。 そのために、転写段階や翻訳段階において、精密な調整が行われています。 この部分に、L7とL12と書かれている遺伝子があります。 リボソーム研究の初期において、各リボソームタンパクは、分子量の違いにより電気泳動により分離されました。 また当時は,ペプチドを構成するアミノ酸配列の決定が非常に困難な時代でした。
このL7-L12遺伝子の産物であるタンパクは、そのN末端が修飾を受けている場合と受けていない,2通りがあることが細胞内に存在することが知られています。 リボゾムタンパクを個々のタンパクとして電気泳動で分離した場合、修飾されたタンパクと修飾されていない産物は、異なる分子量を持つタンパクとして分離されます。 しかし、この2つのタンパクをコードする遺伝子は同じで、N末端のアミノ酸の修飾の有無の違いしかありません。タンパクの名前とするとN 末端のアミノ酸に修飾がある場合がL7,それが無い場合はL12というふうに2つの名前を持っているのは,このような事情によります。
リボソーム粒子には各タンパクが基本的には1つずつ入ることで,全体が構成されるのですが、このL7、L12については、Large subunit内に4個あるということが知られています。 L7とL12のタンパクはダイマーを作り、そのダイマーが2つLarge subunitの中にありますので、計4分子存在します。
リボソームタンパクのネーミングですけれども、基本的にはその分子量の大きいものから番号を振られていきます。Large subunitを構成するタンパクですとL1、L2、L3というふうにナンバリングがされます。Small subunitの中で一番分子量の大きいものをS1、次をS2というふうなナンバリングが行われます。
12. Saccharomyces cerevisiaeにおけるribosomal RNAの転写ユニット
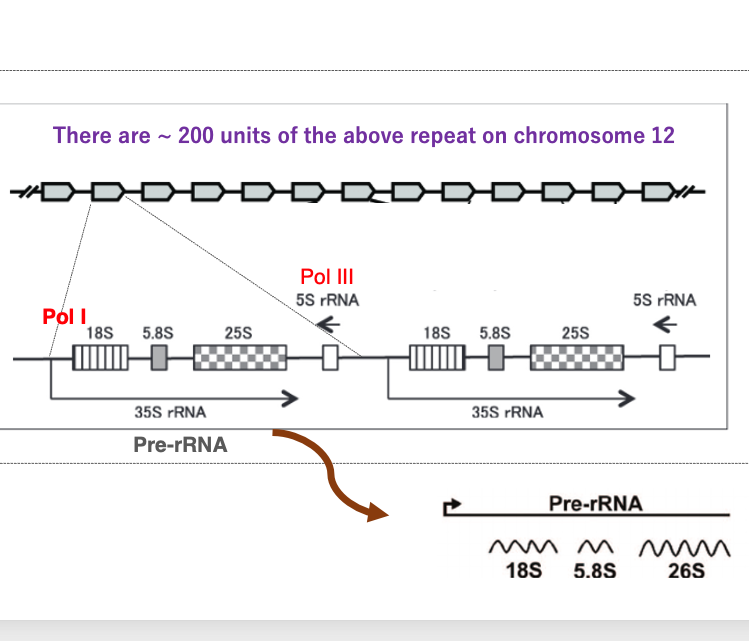
次に、真核生物のribosomal RNAの転写について、酵母のSaccharomyces cerevisiaeを例にして解説をしていきます。 Saccharomyces cerevisiaeでは、ribosomal RNAをコードする転写ユニットが 12番染色体上に200回,繰り返して配置されています。
これが1つの転写ユニットを示しています。 1つの転写ユニットの中には、18S ribosomal RNA、5.8S ribosomal RNA、25S ribosomal RNA、5S ribosomal RNA遺伝子が含まれています。18S、5.8S、25S ribosomal RNA遺伝子をPolymerase 1がこの向きに転写します。その転写分は35S rRNAと呼ばれます。
一方,5S ribosomal RNAはPolymerase 3によってこの向きに転写されます。 Polymerase 1で転写された35S ribosomal RNAは、 その後、RNaseによって切断され、最終産物である18S、5.8S、26S ribosomal RNAになります。
13. 核小体におけるリボソーム粒子の組み立て
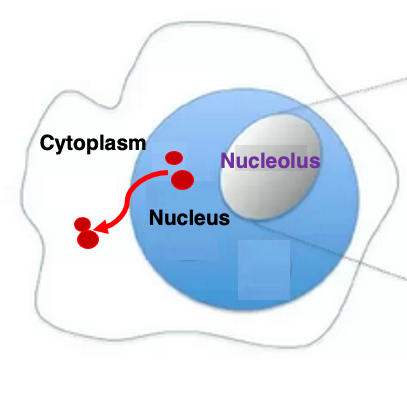
真核生物の場合、ribosomal RNAをコードする遺伝子は、核の中にある核小体の領域で転写されます。
4種類のribosomal RNA が加工されて出来上がりますと、細胞質からリボゾームタンパクが核内に移送されて、立体構造を作りつつあるribosomal RNAの上に順次付着していくものと考えられています。
途中まで出来上がったリボゾーム粒子は、核小体から核膜を通って細胞質の中に輸送され、そこで最終的なファインチューニングがされて組み立てが終わるというふうに考えられています。 したがって、核小体はリボゾーム粒子の組み立て場所であると言われています。
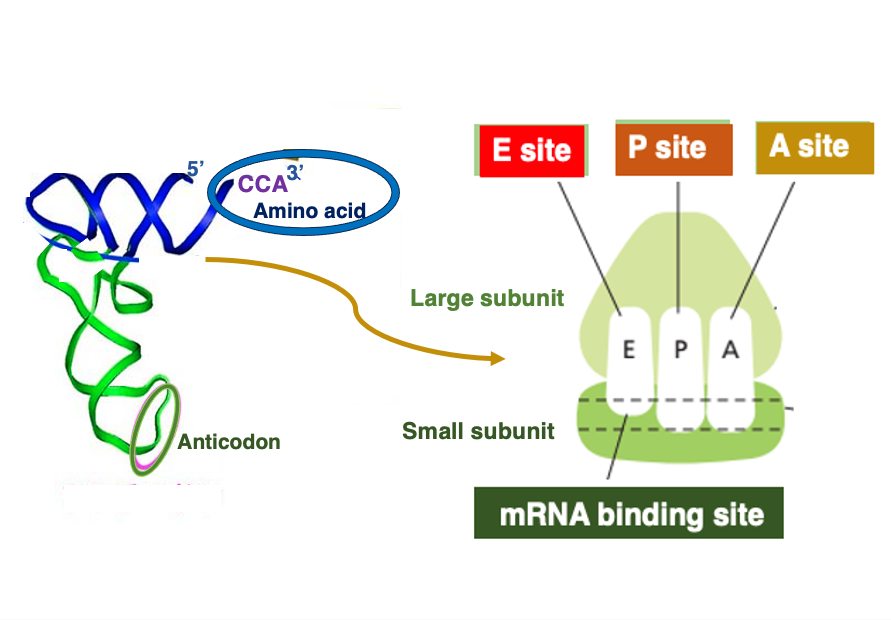
14. リボソームにある内部空間のイメージ
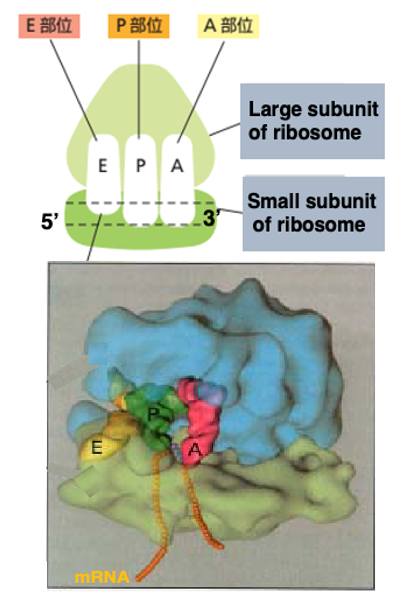
この図はリボソームにある内部空間のイメージ図です。真核生物の場合でも、原学生物の場合でも、リボソームの中には3つの大きな空間があります。 それぞれの空間は、1つのtransfer RNAがちょうど入れるぐらいの大きさを持っています。
mRNAの5 prime 方向から3 prime 方向の順に,Eサイト、Pサイト、Aサイトと名前が付けられています。 EサイトはExit siteの略で ペプチド鎖を放出した tRNAがリボゾームから離脱していくサイトです。 Pサイトには,合成途中のペプチド鎖を3 ‘末端に持っているtRNAが入るサイトです。
Aは、アミノ酸を結合させたtRNAが入るサイトです。 これが,そのイメージ図です。Large subunitとSmall subunitがありますが、Small subunitはmessenger RNAをこのように捕まえています。
15. ペプチド鎖の伸長反応
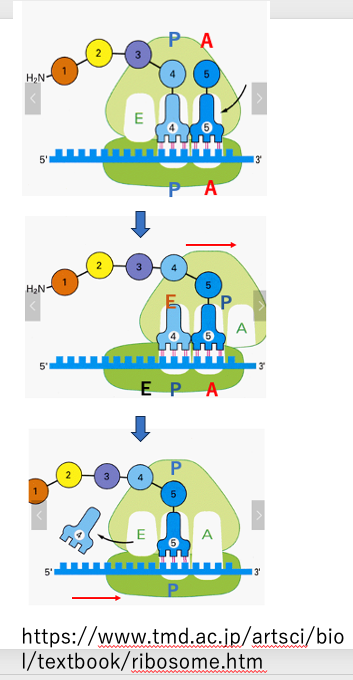
ペプチド鎖の伸長反応を行っている途中のリボソームの動きと transfer RNAの動きについて説明をします。この図では,Pサイトに 4つのアミノ酸がつながったペプチドを持つtransfer RNAが入っています。 その右側には5番目のアミノ酸を連れた transfer RNAが侵入しようとしています。この5番目のtransfer RNAについて メッセンジャーRNA上のコドンと transfer RNAのアンチコドンの対合正しいということが確認されると, リボソームのLarge subunitは 3塩基ぶん,3 prime方向にズレます。それと同時に ,4番目のtransfer RNAに付着していたペプチド鎖は 5番目のトランスフォアRNAが運んできたアミノ酸と結合します。つまり,ペプチド鎖の転移が起きます。ペプチド転移終了直後のtransfer RNAの位置を見ますと, 4番目のアミノ酸を運んでいたtransfer RNAは, Large subunitのサイトで見ればEサイトですが ,small subunitは動いていませんので small subunitではPサイトに入っていることになります。 またペプチド鎖を受け取ったtransfer RNAは,Large subunit のPサイト,Small subunit のAサイトでつくられたcavityに位置しています。このように,transfer RNAはLarge subunitとsmall subunitでは,異なるサイトによって構成されている混成サイトに収まっています。
次にsmall subunit 3塩基分,3 prime 方向にずれます。この時,アミノ酸を放出しているnumber 4 transfer RNAは Eサイトから離脱して,細胞質内に移行します。small subunit 3塩基分,3 prime 方向にずれた為,ペプチド鎖を持っている number 5 transfer RNAは,Large subunitからしてもsmall subunitからしても Pサイトで構成されたcavity内に収まっています。またその右側には、正規のA siteがありますが,このcavity内にtransfer RNAは入っていませんので,ここにアミノ酸を付着されたnumber 6 transfer RNAが侵入してくることができます。
number 6 transfer RNAのアンチコドンがmessenger RNA上のコドンとの間で正しい塩基対合が成立することが確認されると,再びペプチド鎖の転移がおこります。このような一連の反応の繰り返しにより,ペプチド鎖が1アミノ酸残基ずつ伸びていきます。
16. ペプチド鎖伸長の化学反応
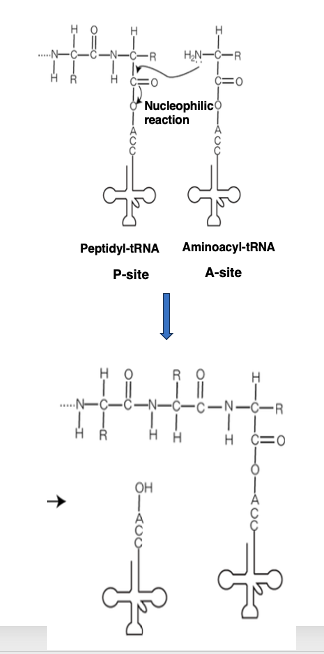
リボソームで行われているペプチド鎖の伸長反応を有機化学反応的に見ると, このように現されます。
今、P-siteに位置している,このtransfer RNAはペプチド鎖をその3 prime末端に 付けています。 一方,Aサイトには アミノ酸を付着したアミノアシルtransfer RNAが 入っています。
このtransfer RNAのアミノ基が ペプチド鎖のカルボキシに対して求核攻撃を行うということで,一アミノ酸残基,伸長したペプチドが形成されます。
17. Peptidyl Transferase Center: 23S ribosomal RNAが持つ酵素活性
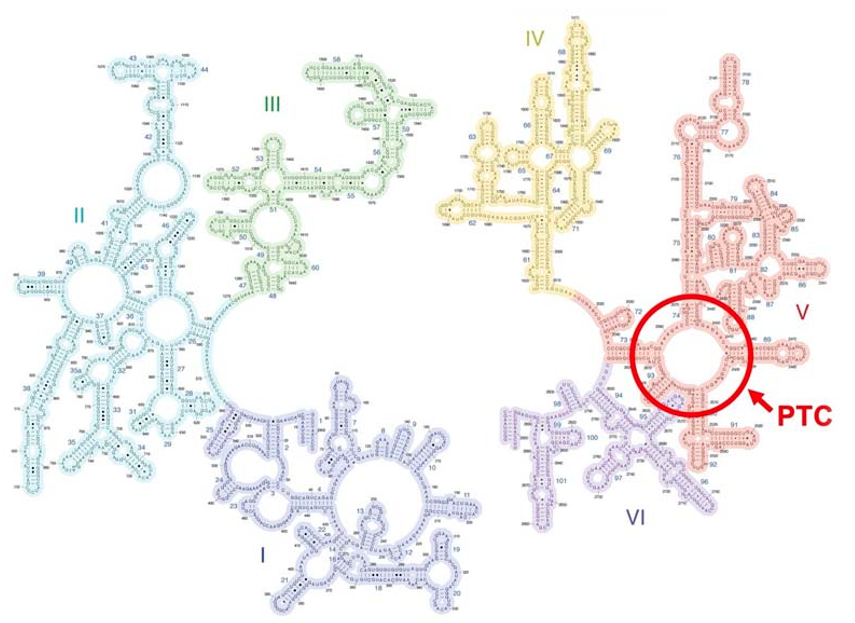
今説明しました ペプチド鎖の転移反応は,リボソームが持つ 触媒活性によって,常温でも 非常に速やかに行われます。 その触媒活性を持っているのはLarge subunitの中にある,23S ribosomal RNA のこの部分にあります。この部分はPeptidyl Transferase Centerと呼ばれます。
つまり23S ribosomal RNAは 触媒活性を持つ生体分子だということができます。触媒活性を持っている生体分子の多くはタンパクですが,このように, 生体反応において触媒活性を持っているRNA分子が存在することが知られてきました。そこで,RNAであって, 触媒活性を持つ酵素として働くようなものを特にリボザイムと呼んでいます。
ribosomal RNA以外の代表的なリボザイムとしては,spliceosomal RNAがあります。細胞の中で,タンパクとRNAの複合体が複雑な生化学反応を行う場合,活性中心はタンパクではなくRNA側にあります。
これは,生命進化の初期において酵素活性が持っていたのはタンパクではなくRNA分子であり,タンパクはRNA分子が酵素活性を発揮するために必要な立体構造を安定的にとることを介助する分子であったことに起因しています。
これでこの講義を終了します。
コメント