
この講義の内容は下記のYouTubeサイト(再生リスト:生命科学を専攻する学生の為の分子生物学講義)で
https://youtu.be/fkbzG1EPY9w
動画として視聴できます
1. 講義内容

この講義は下記のような内容を含んでいます。
1. Barbara McClintockによるゲノム上を動く転移因子の必要性の提唱
2. ヒトゲノムにおいる様々なトランスポゾンの比率
3. トランスポゾンの分類:DNA transposon,Retrotransposon
4. Retrotransposon内の分類:LTR型RetrotransposonとNon- LTR型Retrotransposon (LINE, SINE)
5. DNA transposonの転移様式
6. LTR型Retrotransposonの転移様式
7. LINEの転移様式
8. SINEの転移様式
9. Transposon転移抑制メカニズム
Key words: Barbara McClintock, DNA transposon, Retrotransposon, LTR- Retrotransposon, Non-LTR Retrotransposon, LINE, SINE, transposase, gag, pol, env, integrase, VLP, telomerase, copy and paste, target-primed reverse transcription, endonuclease
2. ゲノム上を動く転移因子の必要性

メンデルは花の色の違いなどを観察して,ゲノム上に形質を決定する遺伝子を仮定しました。それにより,掛け合わせによって生じた花の色の分離比をうまく説明できることを発見し,1865年に論文を発表しました。左側に示した赤い花と白い花の分離比などはその例です。メンデルが仮定したメンデル遺伝子は,染色体上に安定に配置されており,かつ常に発現されている事になります。
それから,およそ90年後の1951年に,Barbara McClintockはトウモロコシの遺伝子地図の作成実験を通じて,ゲノム内には転移する遺伝的な因子がなくては説明がつかない現象がある事を発見し,そのような転移性の因子をtransposon (トランスポゾン)と名づけました。
図に示したトウモロコシでは赤紫の種子と黄色の種子が混ざっていますが,このような種子の色の変化は,メンデル遺伝では説明できない事を発見しました。McClintocが提唱した染色体上を転移する転移因子の存在は,その後の分子生物学の発展によりDNA配列レベルで確認され,彼女は1983年にノーベル賞を受賞しています。
今では,全ゲノム配列のデータ解析からトウモロコシのゲノムの80%は,トランスポゾンやその残骸から構成されていることが分かっています。
3. アサガオに見られるトランスポゾン転移による変異

アサガオ(Japanese morning glory)には様々な変異体が見られます。これらの変異体の多くも,ゲノムDNA複製の際に起こった塩基突然変異ではなく,トランスポゾンがゲノム上を転移し,そのコピー数を増やすことで既存の遺伝子を破壊したり,その発現に変化を及ぼしたことが原因です。
このようなトランスポゾンの振る舞いは人間から観ると,ひたすら自分のコピーを増やすことが生存目的の利己的な因子のように思えます。この講義では,トランスポゾンの転移様式とコピー数が増加の機構ついて解説します。
4. ヒトゲノムにおいる様々なトランスポゾンの比率
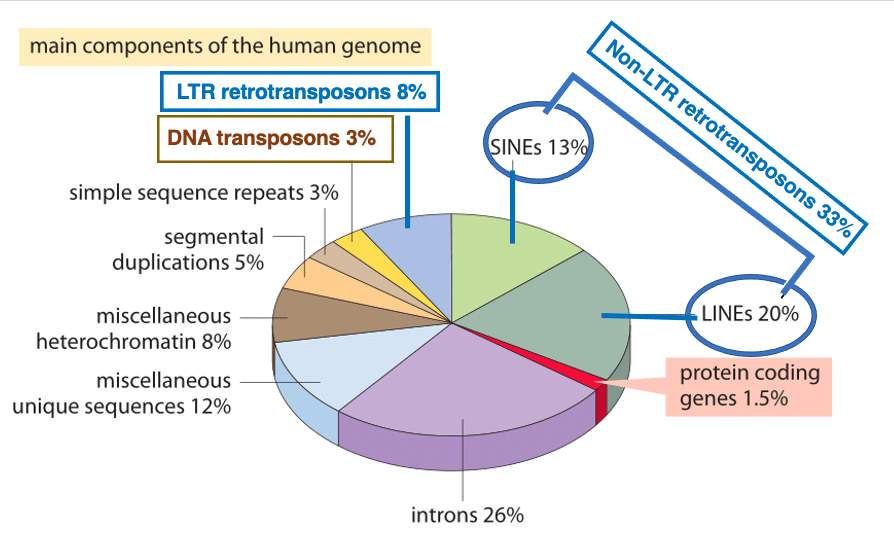
この円グラフはヒトのゲノムを構成している要素の比率を示しています。まずタンパクをコードしている遺伝子のエクソン部分を全部合わせると1.5%, 一方で,イントロンを合計すると26%あります。例外的にイントロンを持たない遺伝子として,ヒストンやG-タンパク,一部の転写因子がありますが,それ以外のタンパク遺伝子は複数のイントロンを持っています。
トランスポゾンには,DNAトランスポゾンと,レトロトランスポゾンがあり,レトロトランスボゾンは更に,その両末端に200-600 bpの Long Terminal Repat (LTR)を持つ型とNon-LRTに分かれます。DNAトランスポゾンは,ゲノムの3%, レトロトランスポゾンの一種であるLTR型が8%,Non-LRT型は更にSINEとLINEに分類されますが,Non-LRT型が33%もあります。DNAトランスポゾンの3%とレトロトランスポゾンの41%を合計すると,トランスポゾンはゲノムの約半分,44%を占めています。
しかし,多くのトランスポゾンは現在では転移活性を失った状態にあります。ヒトでは,転移活性を持つDNAトランスポゾンは,現在存在していません。また,activeなretrotransposonは,LINE-1, Alu, SVAの3種類に限られています。
5. 様々な生物種においてトランスポゾがゲノムに占める割合

これは,様々な生物種においてトランスポゾンがゲノムに占める割合を円グラフで示しています。このように,生物種において,トランスポゾンの比率は大きく異なります。ヒトは,どちらかと言えばトランスポゾンの比率が大きい生物と言えます。
Drosophila melanogasterでは,3-5 %であるのにたいして,Drosophila simulansでは11-13%なので,近縁種であってもトランスポゾンの存在比は大きくことなることが分かります。
Activeなトランスポゾンがどのようにして生まれ,inactiveになっていくのかは興味深いテーマだと思います。少なくても,一部のトランスポゾンは種間を超えて転移していると考えられています。
6. DNA transposonとRetrotransposon
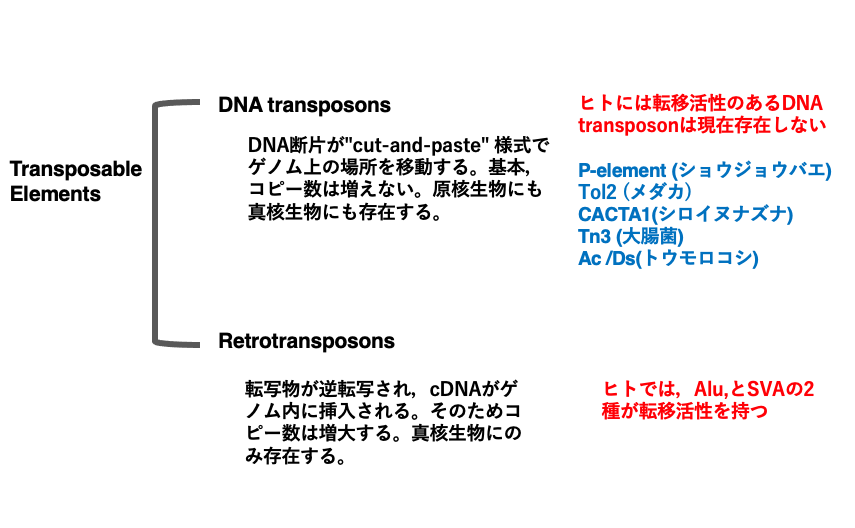
トランスポゾンはその転移様式から,DNA transposonとRetrotransposonの2つに大別されます。DNA transposonは,DNAである転移因子がゲノムから切り出されて,cut-and pasteで別の場所に挿入されます。cut-and paste形式での転移ですので,ゲノム内でそのコピー数は,増え難いと思われます。DNA transposonはDNA virusと共通の祖先を持っていると考えたれています。
現在,ヒトには転移活性のあるDNA transposonは存在しませんが,かつて,転移活性を持っていたと思われる残骸がゲノムの3%を占めています。一方,ヒト以外の真核生物や原核生物では,転移活性のあるDNA transposonが知られています。
Retrotransposonは,RNAを介して,そのコピー数が増えるトランスポゾンです。転写されたmRNAが逆転写酵素によってDNAに変換されたcDNAがゲノムに組み込まれます。転写分子であるmRNAは複数コピー生産されるので,多数のcDNA分子が作られる可能性が高く,コピー数が増えやすいという特徴を持っています。Retrotransposonは真核生物にのみ存在し,原核生物には存在しません。
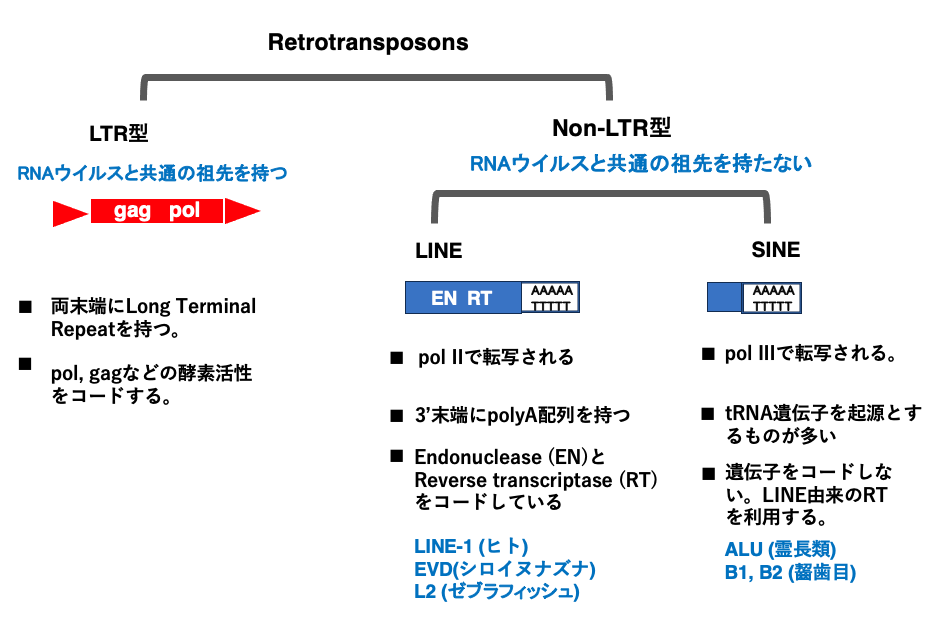
7. Retrotransposonの分類
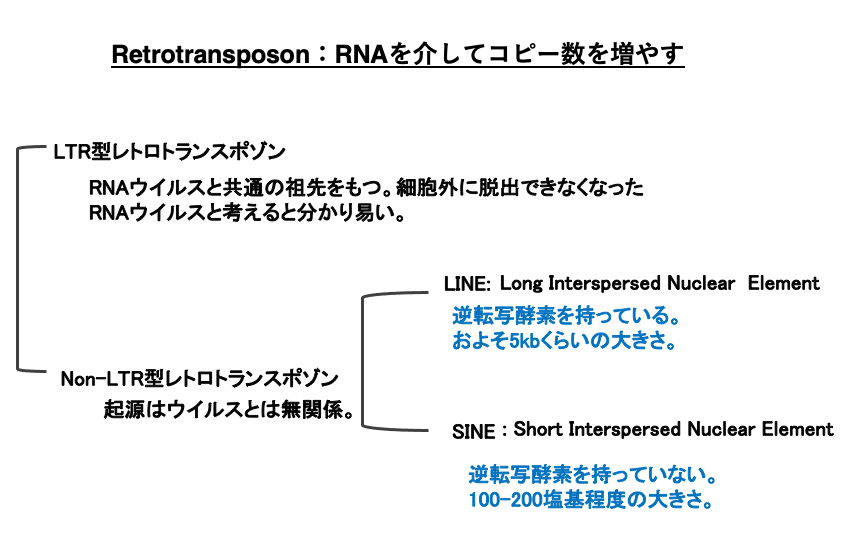
Retrotransposonは,LTR型retrotransposonとNon- LTR型retrotransposonに大別されます。LTR型retrotransposonはReverse transcriptaseを介して増殖するRNAウイルスと共通の祖先を持っていると考えられています。従って,RNA virus型retrotransposonともいえます。
一方,Non- LTR型retrotransposonは,RNAウイルスとは祖先の共通性がありません。Non- LTR型retrotransposonには,5,000塩基対程度の大きさを持ち,Reverse transcriptaseを内部にコードしているLong Interspersed Nuclear Element (LINE)と,100-200塩基程度の大きで逆転写酵素をコードしていない,Short Interspersed Nuclear Element (SINE)の2つに分かれます。
8. DNA transposonの転移様式
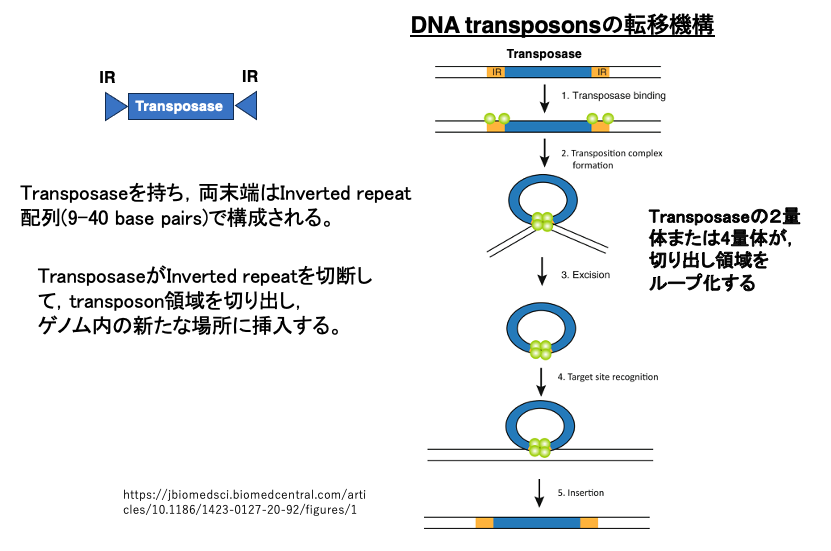
まず,DNA transposonの転移様式について解説します。このトランスポゾンの両端には,9-40塩基のInverted repeat配列があります。このInverted repeat配列に挟まれた領域にはTransposaseがコードされています。
TransposaseはInverted repeat配列に付着して,両末端にあるInverted repeat配列を手繰り寄せて,ループ化して切断します。切断されたDNA transposonとTransposaseの複合体が,新たなゲノムのサイトを切断して,切り出したDNA断片を,その場所に挿入します。DNA transposonが遺伝子内に挿入された場合は,遺伝子の破壊が起こります。また,イントロン内に挿入されたような場合でも,正常なスプライシングを妨げる可能性があります。
9. 大腸菌にあるDNA transposon Tn3
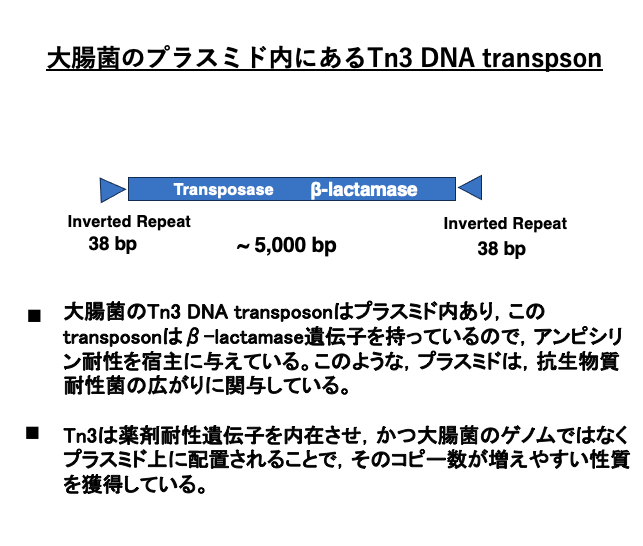
大腸菌にあるDNA transposon Tn3は5,000塩基弱のトランスポゾンです。その両末端には,38塩基対のinverted repeatがあります。内部にはTransposaseの他に,β-lactamase遺伝子がコードされています。そのため,抗生剤であるアンピシリン等を分解できるので,宿主がこのトランスポゾンを持っているとアンピシリン耐性となります。
更に,Tn3はゲノムではなくプラスミド内に挿入されているので,プラスミドが他の菌体に受け継がれることで,アンピシリン耐性が広がっていくことになります。視点を変えて言えば,Tn3は薬剤耐性遺伝子を内在させプラスミド内に配置されるのは,自分自身のコピー数を増やすのに有利な条件を獲得していると言えます。
10. Retrotransposonの分類
次にRetrotransposonについて説明をします。Retrotransposonでは,自身をコードする領域がRNA polymeraseによって転写され,それがReverse transcriptaseにより逆転写されます。cDNAは,染色体の新たな場所に挿入されます。従って,多くの転写物が生産されれば,結果的に複数のcDNA分子が生まれるので,ゲノム内のコピー数は急激に増えることになります。つまりRNAを介して増えるtransposonと言えます。
Retrotransposonで,両末端にLong Terminal Repat (LTR)を持つLTR 型では,RNAウイルスと共通の起源を持ち,細胞に侵入したRNAウイルスの中で,細胞外に脱出するために必要なenvelope殻タンパク遺伝子を喪失したRNAウイルスだと考えると分かり易いと思います。
一方でNon-LTR型は,その起源がRNAウイルスとは関係性がないと考えられています。Non-LTR型のretrotransposonは更に,LINEとSINEに分類されます。LINEとSINEは同じ転移機構を持ちますが,その起源となった分子には共通性がありません。LINEは分子内には,RNA polymerase IIで転写され,その内部には転移に必要なタンパクがコードされますが,SINEにはそれがありません。SINEはtRNAや5S rRNAのようなRNA polymerase IIIで転写される小さなRNA遺伝子を起源としています。
LNEもSINEもその3‘領域のDNA配列内に20-50塩基対の3’-AAAA-5’/5’-TTTTT-3’ DNA配列を持っています。従って,それらの転写物はpoly A RNA配列をその3’末端に持っています。このpoly A配列は,Non-LTR retrotransposonが逆転写され,新たなゲノムの位置に挿入さるために,重要な役割を持っています。
11. LTR を持つRetrotransposonの転移様式

LTR を持つRetrotransposonの転移様式について解説します。このRetrotransposoはRNA virusと共通の祖先を持っています。
12. Reverse transcriptaseで増えるretrovirusのゲノム構造
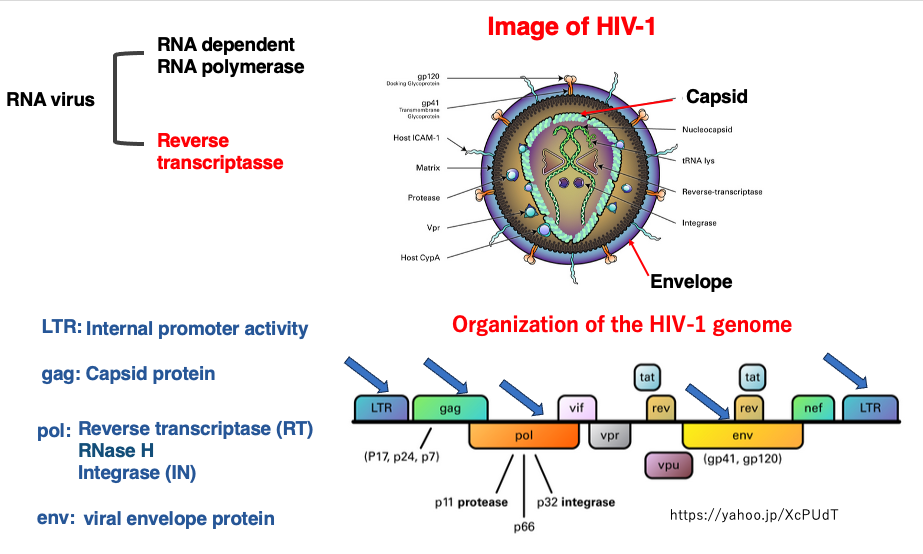
LTR型のretrotransposonは,ウイルスと共通の祖先をもつので,まずは,Reverse transcriptaseを介して増えるretrovirusについて復習しておきます。これは,ヒトHIV-1の外観とゲノム構成の図です。
この種のRNA virusは,両末端に長いinverted repeat (LTR)をもち,その配列はpol IIに対するpromoterとして作用します。また内部にはgag pol, envという3つの遺伝子を共通に持っています。Gag遺伝子には,capsidタンパクがコードされています。Pol遺伝子の産物は,proteinaseで切断されて,Reverse transcriptase, RNaseHとIntegrase活性を持つタンパクとなります。Env遺伝子には,ウイルスの最外層の殻タンパクであるenvelopeタンパクがコードされています。
13. LTR retrotransposonのゲノム構成
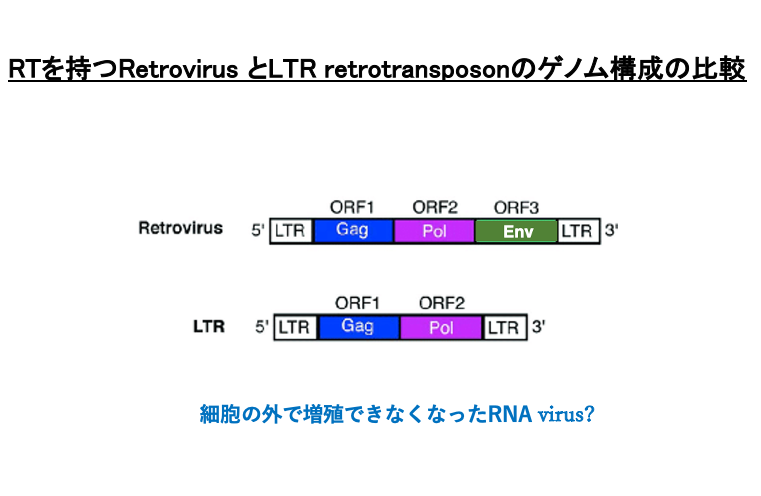
これは,Reverse transcriptaseを持つRetrovirus とLTR retrotransposonのゲノム構成の比較です。LTRをもつ transposonでは, retrovirusが持っていたenvelopeタンパクをつくる遺伝子を持っていません。
LTR retrotransposonは,ゲノム内に侵入したRetrovirusが,細胞に潜伏中にenvelope遺伝子を欠損して,細胞外で増殖する能力を欠損することで生じた分子であると考えると分かり易いと思います。
14. LTRを持つ retrotransposonの転写と転移の概略
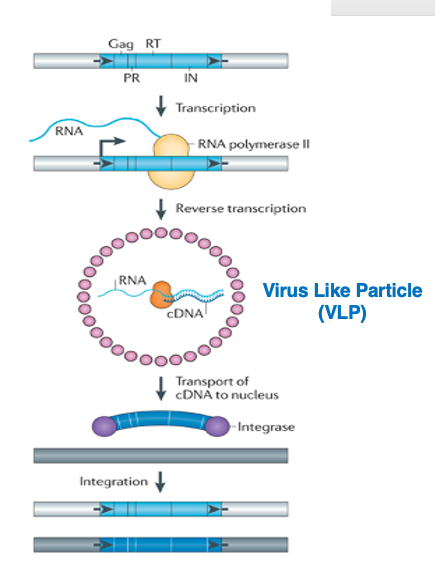
LTRを持つ retrotransposonの転写と転移の概略について説明します。
転写は5’-側のLong Terminal Repeat内にあるpromoterから,RNA polymerase IIによって行われます。mRNAは細胞質に移行して宿主のリボソームにより翻訳されます。gag遺伝子にコードされていたcapsidタンパクが集合して形成された粒子内には,mRNA, pol遺伝子由来のReverse transcriptaseとIntegraseなどが取り込まれてVirus Like Particle (VLP)が形成されます。この中で,逆転写が行なわれ,最終的には2本鎖cDNAが作られます。
2本鎖cDNAの末端にintegraseが付着した状態で,cDNAは核内に移行します。Integraseの作用によって,cDNAはゲノム内に挿入されます。挿入箇所については,ほぼランダムであると思われています。遺伝子内に挿入されれば,遺伝子破壊を引き起こします。また,両末端にあるLTRは,RNA polymerase IIのpromoter活性を持っているので,近傍の宿主遺伝子の転写に影響を与える場合があります。
このmRNAの逆転写の過程はかなり複雑です。5’-UTRの途中から3’-UTRの末端までがmRNAとして作られますが,3’-UTR配列を利用することで,形成されたcDNAには元と同じ完全長のLTRが5’末端と3’末端に配置されたcDNAが作られます。このため, 転移したコピーDNAでもpromoter活性が維持されているので,ゲノムのどこに挿入されても,自身の遺伝子を転写することができまるactiveなトランスポゾンのコピーとなっています。
15. LTRを持つ retrotransposon転移の詳細なメカニズム
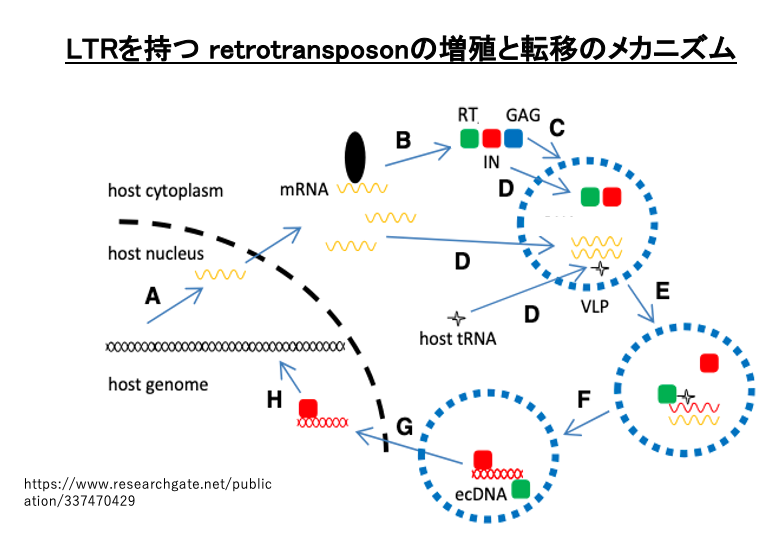
別の図でLTRを持つ retrotransposonの転移を復習します。核内でトランスポゾンがpol IIによりLTRから転写されます。細胞質に移行したmRNAは翻訳さて,gag領域からはcapsidタンパク(青色)が, pol領域からはReverse transcriptase(緑色)とintegrase (赤色)が作られます。capsidタンパクが集合して,Virus Like Particle (VLP)が形成されます。この粒子のなかでmRNAが逆転写され,最終的には2本鎖cDNAとなります。この逆転写反応のprimerとして,宿主のtRNAが用いられます。
cDNAとintegraseは,核内に移行します。Integraseによって,cDNAはゲノム内に挿入されます。このLTR-retrotransposonのcDNAは,プロモーター配列を含む損傷のない5’-LTRを保持しているので,更に転移することができます。
16. LINEの転移機構
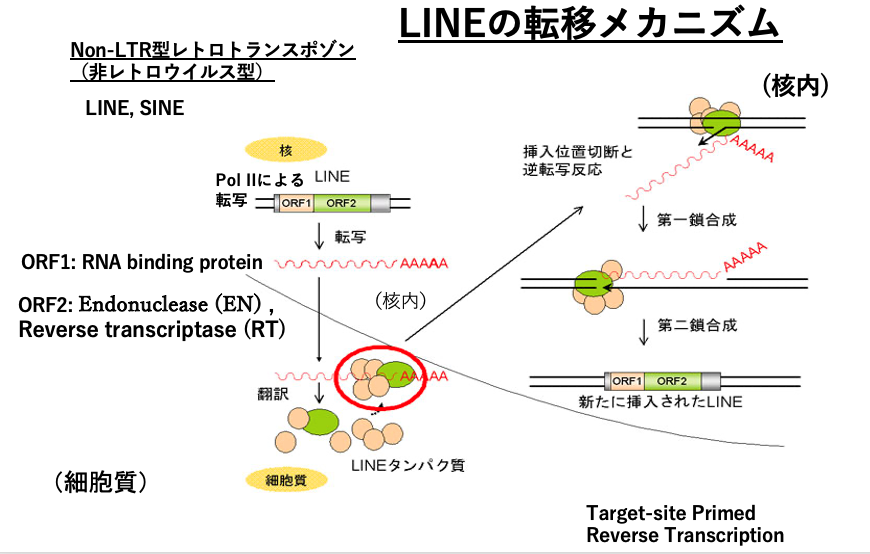
次にNon-LTR retrotransposonの一種,LINEの転移について解説します。さきほどのLTR retrotransposonはRNA virusと近縁関係にありましたが,Non-LTR retrotransposonもRNAの逆転写を介して自己のコピーを増やしますが,RNA virusとは,reverse transcriptaseを除き共通の祖先を持っていません。
Non-LTR retrotransposonはORF1とORF2をコードする2つの遺伝子を持っています。ORF1は自身のmRNAに付着するRNA binging proteinをコードしています。ORF2には,弱い特異性を持つendonuclease (EN)とReverse transcriptase (RT)がコードされています。
転写は,5’-UTR内にあるプロモーターからRNA polymerase II によって行われ,宿主のリボソームにより翻訳されます。翻訳産物であるRNA binging protein,endonuclease,Reverse transcriptaseが自身のmRNAに付着して,mRNAは核内に戻されます。
核内で,Endonuclease によりゲノムが切断されると,そのDNA切断点の3’-OHからretrotransposon mRNAを鋳型とした逆転写が始まります。最終的に2本鎖化されたDNAが,ゲノムの切断点に挿入されます。このような機構はtarget-primed reverse transcription transposition と呼ばれます。
17. LINE-1転移機構の詳細
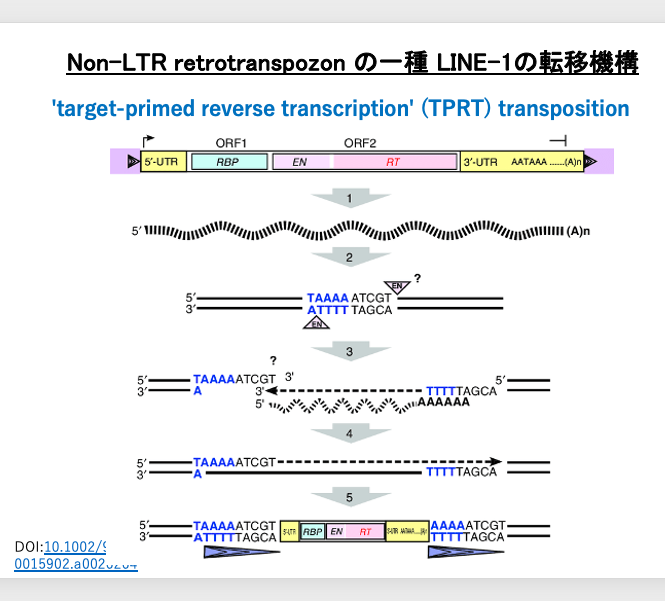
別の図でLINE-1のtarget-primed reverse transcription transpositionをもう少し詳しくみてみましょう。5’-UTR領域のプロモーターからRNA polymerase IIによって,転写が起こり,RNA binding proteinをコードするORF1, endonucleaseとRTコードするORF2及びpoly A配列を含む3’-UTRまで転写されます。
通常,RNA polymerase IIは,promoterではTATA BOXの20-30塩基下流から転写が起こります。従って,pol IIで転写されたmRNAの内部にはプロモーター配列は含まれていません。一方,LINEのpol II promoterは特殊です。Internal promoterの一種でpromoter配列の上流から転写が起こるので,そのmRNAにはプロモーター配列が含まれています。従って,そのcDNAにもプロモータが含まれているので,転移後のコピー分子であっても,再び自身の遺伝子を転写することができます。
LINE-1のendonucleaseは弱い塩基特性を持っており,5’-TTTA-3’を認識してTとAの間を切断します (5’-TTT|A-3’)。認識配列には制限酵素のような対象性が必要性ないので,もう一方の鎖の切断点は不明確ですが,5’-TTT|A-3’配列から5塩基くらい上流の位置ではないかと思われています。この図では切断されたDNAは9塩基の3’突出配列となっています。転写されたLINE-1の3‘-末端のpoly Aが切断されたゲノムDNA 3’突出末端にあるTTTと水素結合を生成します。付着したLINE-1 mRNAを鋳型として,DNA 3’が伸長する形で,逆転写が起こります。最終的にはLINE-1 mRNAが完全な2本鎖DNAとなります。この時,9塩基の3’突出として切断されていたゲノムの突出部も2本鎖に修復されます。そのため,LINE-1のコピー分子の両末端は,9 bpの順方向のrepeat配列に挟まれています(図の細長い三角形)。
18. SINEの転移メカニズム
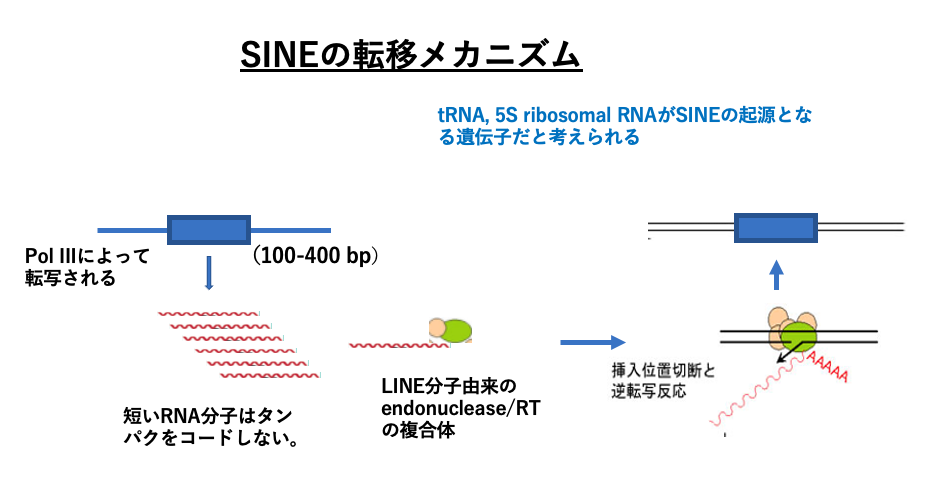
次にLINEと同じくLong terminal repeatを持たないretrotransposonの一種ですが,もっと短い分子であるSINEの転移メカニズムを解説します。
SINEはRNA polymerase IIIで転写されるtRNAや5S rRNA遺伝子を起源とする分子が大半です。そのため100-400 bpと短くまた内部にはORFを持ちませんが,起源のtRNAや5S rRNAとことなり,遺伝子のDNA3‘末端に短いpoly A配列があります(5’-AAAAA-3’/3’TTTTT-5’)。この点はLINEと同じです。
SINEの転写物に,細胞内のLINE由来のendonucleaseやReverse transcriptaseが付加し,核内に移行します。LINE由来の酵素を借用して,LINEと同じ’target-primed reverse transcription’ (TPRT) transpositionにより,自己のコピーをゲノム内の新たな位置に挿入します。LINEの酵素を借用して自己のコピーをゲノム内につくるという点においては,SINEはLINEよりも寄生性が強い分子と言えます。
19. SINEの転写に使われるRNA polymerase IIIのpromoterの特徴
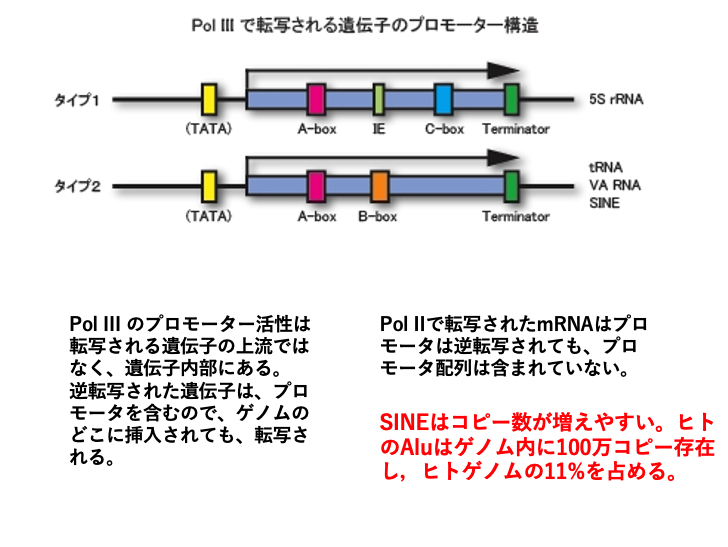
この図に示したように,RNA polymerase IIIのpromoterはそれが転写する遺伝子に含まれています。そのため,転写物がDNA化しても,再び転写されます。また,短い分子なのでLINEとは異なり,逆転写が途中で止まってしまうことが少ないと考えられます。
そのため,増殖しやすい分子であると考えられます。実際,ヒトでactiveなSINE分子の一種のAluは120万コピー存在します。
20. ヒトゲノムにあるtransposonのコピー数
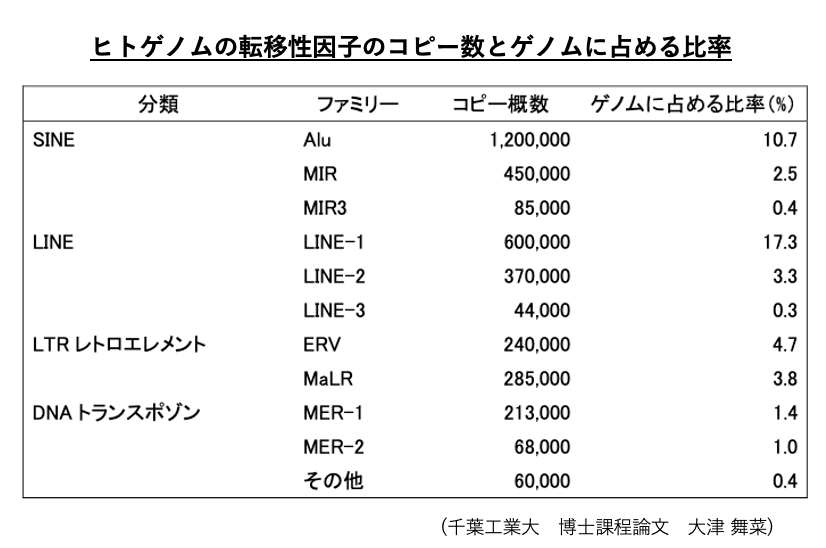
もう少し,詳しくヒトゲノム内にあるtransposonのコピー数をみると,この表のようになります。
ヒトでは,コピー数の多い順にSINE, LINE, LTR-retrotransposon, DNA transposonとなっています。
21. Reverse transcriptaseとTelomeraseの近縁性
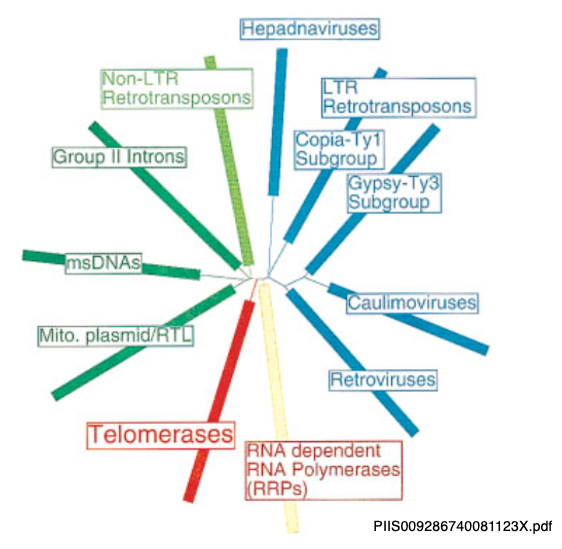
LTR型であれNon-LTR型であり,Retrotransposonの転移にはreverse transcriptaseが必須です。Reverse transcriptaseは,RetrovirusやGroup II intronの転移にも必須な酵素です。
そればかりではなく,ゲノム末端の反復配列に関わるテロメラーゼもreverse transcriptaseの一種です。これらの系統進化関係は不明ですが,テロメラーゼは,線状ゲノムを持つ真核生物の誕生に必須な酵素です。それがretrotransposonのReverse transcriptaseと近縁であることは,非常に興味深いと思います。
コメント