この講義の内容は下記のYouTubeサイト(再生リスト:生命科学を専攻する学生の為の分子生物学講義)で動画として視聴できます
https://youtu.be/q_Jy6vi0wrU
1. この講義は以下の内容について解説をしています

1. 性染色体構成の異常と症状
2. 常染色体トリソミーと症状
3. X染色体のランダムな不活性化
4. 様々なX染色体の遺伝子量補正機構
5. X Inactivation Center (XIC)の発見
6. Xist, Tsix non coding long RNA
7. XICの対合と分離による不均衡なCTCFの分配
8. 不活性化されるX染色体が決定される機構
Keywords : 擬似常染色体領域,PAR, XXY, XXX , XO, 常染色体トリソミー, ターナー症候群, クラインフェルター症候群, メアリー・ライアン(Mary Lyon)の仮説, 三毛猫の体毛色, X inactivation center (XIC), Tsix RNA, Xist RNA, エピジェネティックなヒストン修飾
2. ヒト体細胞の染色体構成
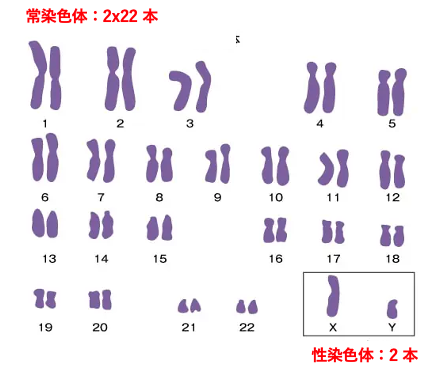
この図はヒトの体細胞の染色体構成を示しています。
ヒトの体細胞は2n (複相)なので,大きい順に1番から22番までの常染色体がペアであります。
また性染色体として,女性の場合はX染色体が2本,男性の場合はX染色体が1本とY染色体が1本ずつあります。
3. X染色体とY染色体の比較
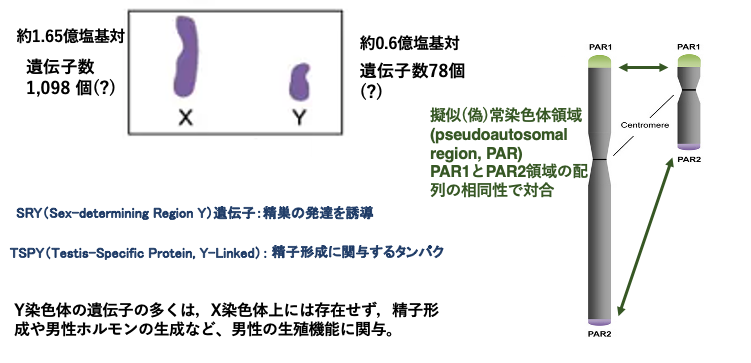
X染色体が約1.65億塩基対で構成されているのに対して,Y染色体は約0.6億塩基対で構成されています。したがって,Y 染色体の大きさはX染色体の3分の1弱の大きさしかありません。
X染色体には1,098個の遺伝子,Y染色体には78個の遺伝子がコードされています。
Y染色体には,精子形成(SRY gene)や男性ホルモンの生成に関する遺伝子(TSPY gene)がコードされています。これらの遺伝子はX染色体上にはありません。
このように,X染色体とY 染色体は大きさも内部の塩基配列も大きく異なっていますが,X染色体とY 染色体は減数分裂時には,常染色体のペア染色体のように対合します。X染色体とY 染色体の対合が可能なのは,2つの染色体で高度に保存された配列を持つ,擬似常染色体領域(pseudoautosomal region, c1とPAR2)がX, Y性染色体の末端にあるためです。
4. 性染色体構成の異常とその影響

受精卵がX 染色体の数について異常を持つ場合,軽度の異常が見られる場合がありますが,日常生活に支障をきたすような異常は現れません。
X染色体を1本のみ持つ女性は,1,000〜2,000人に1人の割合で存在します。この場合,通常の生活に大きな問題はありませんが,身長が低い傾向があります。
X染色体を3本のみ持つ女性も,1,000人に1人程度の割合で存在します。この場合も,身体的な特徴はほとんどありません。
性染色体構成がXXYの男性も1,000人に1人程度の割合で存在します。この場合も,日常生活には問題がありません。
一方,性染色体構成がYのみ,あるいはYYを持つような受精卵は,発生のごく早期で自然流産となるので,このようなヒトは存在しません。これは,X染色体上には,性決定だけでなく生命維持に必要な多くの遺伝子があるため,常染色体とY染色体のみでは,個体を形成するための遺伝情報が揃っていないためです。一方,生命維持に必要な遺伝子がY染色体上には存在しません。したがって,XOやXXの性染色体を持つ個体が生存できます。
これらの事から,常染色体とX染色体があれば,女性を形成するために必要な遺伝子は総て揃っており,女性をベースにして男性的な特徴を持った個体を形成するためにY染色体上の遺伝子が必要だということが分かります。男性は女性をmodifyして作られていることが分かります。体細胞では1本のX染色体が遺伝子を転写できる状態にあり,もう一方のX染色体が高度に凝集して不活性化しているのが正常な状態です。このためXOの個体は,XXの個体と大きな差がありません。また,XXXの個体では3本のうち2本のX染色体が不活性化される機構があるために,大きな異常は見られません。。
5. 常染色体トリソミーの影響
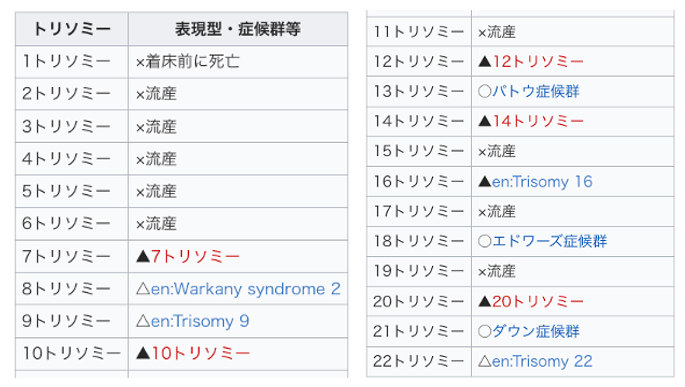
次に特定の常染色体を3本持つ場合(トリソミー)の影響について解説します。
常染色体には大きい順に番号が振られています。
1番から6番のような大きな常染色体がトリソミーとなった場合には,流産が起きている事がわかります。一方,染色体が小さくなると何らかの症候群を発祥していますが,流産にはなっていません。これは,染色体が小さいばあい,コードされている遺伝子の数も限定的であるためだと考えられます。
トリソミーの場合,個体を形成するのに必要な遺伝情報に欠損はありません。トリソミーで様々な異常が引き起こされるのは,複相のゲノムセットを持つ生物で,特定の常染色体が多いと,細胞の中に特定の遺伝子からの代謝物が多くなることで,細胞に有害な影響が現れるからだと考えられます。
常染色体ではX染色体で機能しているような,染色体数の数を把握し,過剰なら不活性化する機構は存在していません。
6. X染色体の不活性化
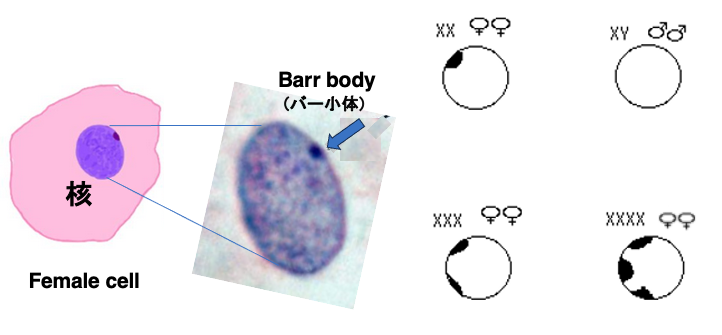
1949年頃に,Barr博士は女性の体細胞のクロモソームを染色すると,細胞周縁部に常に染まる点が観察さることを見出し,Barr bodyと命名されています。Barr bodyは,男性の体細胞では観察されません。このBarr bodyは2つのX染色体の一方が凝集した物であることが分かっています。
XXXの細胞の場合には,Barr bodyは2個,XXXXの場合には3個観察されます。Barr body検査は、1960年代から1990年代にかけ,オリンピックの出場選手に対して使用された性別確認方法の一つでした。
7. メアリー・ライアン(Mary Lyon)の仮説 (1961年)

1961年にメアリー・ライアン(Mary Lyon)は,Barr bodyについて,図に示したような仮説を発表しています。その後,この仮説は正しいことが分子生物学的な研究から証明されています。
8. X染色体のランダムな不活性化
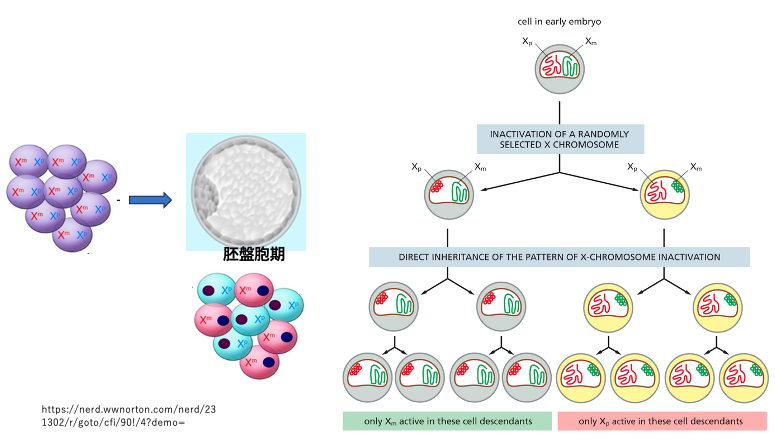
XXの受精卵において,母方由来のX染色体を不活性化するのか,父方由来のX染色体を不活性化するのかは,ランダムに起こります。不活性化されるX染色体の決定は,受精後5日目の胚盤胞期に起こります。また,一旦,不活性化されたX染色体は,その後の体細胞分裂でも不活性化(凝集)が維持されます。
例外的に胎児の生殖細胞の中では,不活性化されていたX染色体の凝集が解かれます。
したがって,次の世代では精子,卵ともにその内部には活性化されたX染色体が入っています。
9. 成人個体のX染色体キメラ
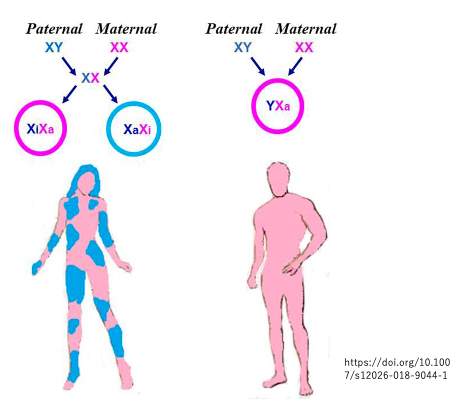
胚盤胞期にランダムに選択されたX染色体の不活性化がその後の発生を通じて維持される結果,成人女性の体では染色体の選択に関して,パッチワーク状の分布が見られます。
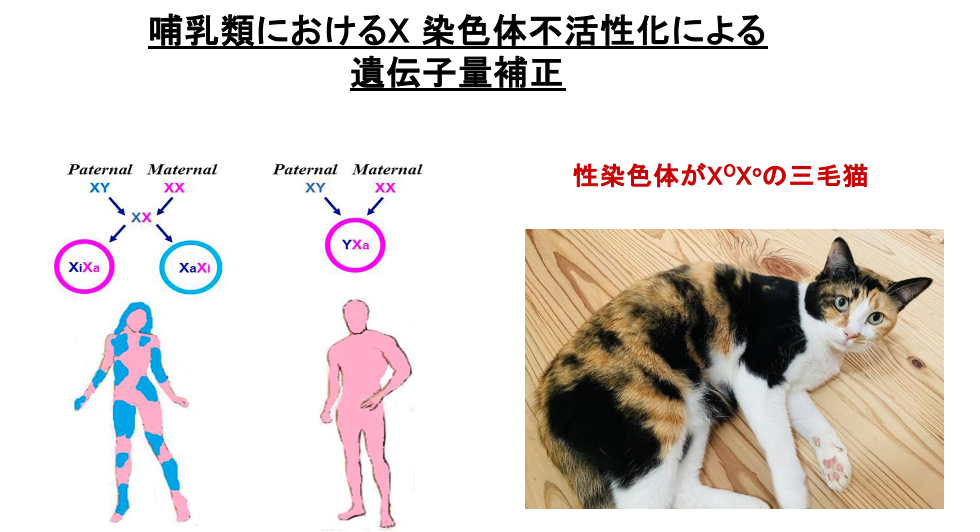
10. 三毛猫の体毛色に関する遺伝子の発現

ネコでは毛の色を決定する遺伝子がX染色体上に配置されています。有性遺伝子のOがあれば,黒色の色素が沈着し,劣性遺伝子のoの場合は茶色の色素が沈着します。一方で,常染色体上の遺伝子Sが発現した領域では,色素の定着が妨げられるため体毛はX染色体上の遺伝子に関わらず白になります。
したがって,三毛猫の性染色体がXOXoの場合,黒い体毛の領域はXOがactiveであり,かつS遺伝子が発現していない,茶色の体毛の領域ではXoがactiveでありS遺伝子が発現していない領域という事になります。白色の領域ではどちらのX染色体がactiveかは不明ですがS染色体が発現している領域という事になります。
11. 様々な生物におけるX染色体の遺伝子量補正のメカニズム
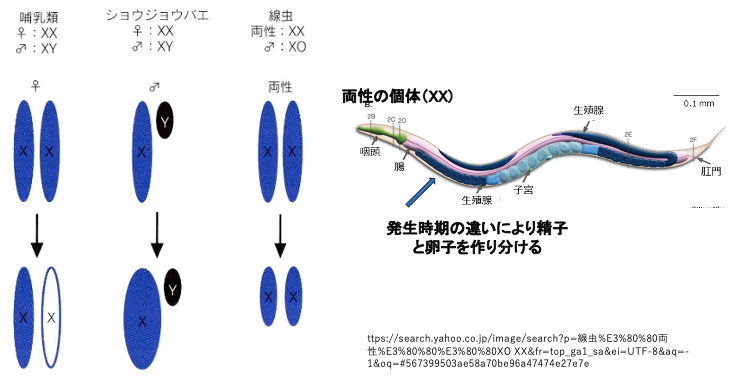
性染色体がXY型の生物種における,性染色体の遺伝子量補正の機構は生物種によって異なります。
哺乳類の場合は,一方のXが完全に不活性化することにより,XYを持つ細胞内でのXの発現量とXXをもつ細胞内におけるXからの転写量が同じになるように補正されています。
一方,ショウジョウバエでは,XYのオスにおいてのみXの発現量が2倍となることでXXであるメスの細胞における発現量と同じになるように補正が行われます。
また,線虫では,XXの個体が精子と卵の両方を生産する両性,XOがオスとなりますが,XXを持つ細胞内では,両方のX染色体の転写量を半分にすることでXO細胞内の転写量と同じになるように補正されています。
12. 不活性化したX染色体でも発現している遺伝子
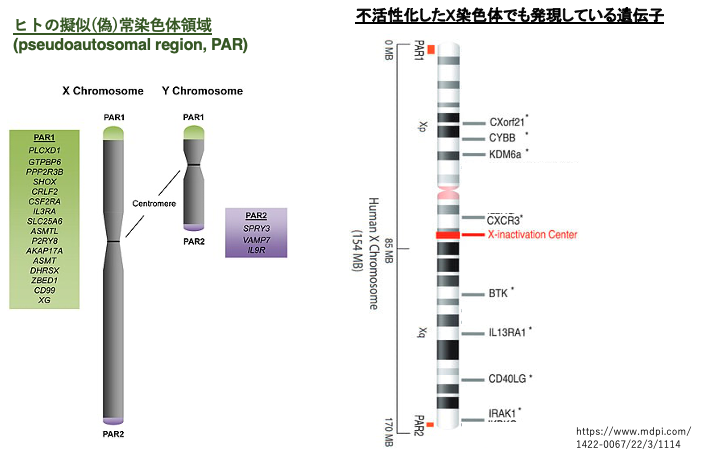
擬似(偽)常染色体領域PAR1とPAR2はX, Y両方の性染色体の両末端にある領域で,この領域が減数分裂時におけるX, Y染色体の対合に使用される事はすでに紹介しました。
この領域にある遺伝子は不活性化したX染色体であっても,不活性化を免れている領域であり,この領域にある遺伝子は発現抑制を受けません。PAR領域は,X染色体でもY染色体でも保存されており,共通の遺伝子がコードされているので, XXでもXYでも,これらの領域にある遺伝子は,常染色体上の遺伝子のように2個の遺伝子が発現していることになります。
ヒトではX染色体において,PAR領域以外の部分にある遺伝子でも不活性化を免れて発現している遺伝子が相当数あることがわかりました(全X染色体遺伝子の約30%)。このような遺伝子で相同遺伝子がY染色体上にない場合,XXでは2個の遺伝子がXYでは1個の遺伝子が発現している事になります。
これが, XO (ターナー症候群)やXXY(クラインフェルター症候群)のような個体において軽度ながら異常が見られる原因となっています。これらの性染色体構成では,XXやXYと不活性化を免れて発現している遺伝子の数が異なっています。
13. X inactivation center (XIC)の発見
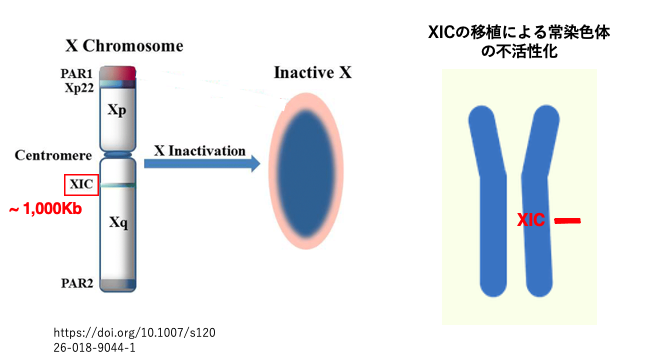
X染色体において特定の領域が欠損した場合,そのX染色体は不活性化しないことが分かり,その領域はXIC (X-inactivation center) 領域と名づけられました。この領域はおよそ1,000 kbからなります。また,この領域を常染色体に移植すると,その常染色体は不活性化することがわかりました。
また,不活性化したX染色体でのみXIC領域から発現している〜17Kbのlong non-coding RNAが検出され,その遺伝子はXist (X Inactive Specific Transcript)と名づけられました。Xist遺伝子は,activeなX染色体ではほとんど発現していませんでした。
14. Xist non-coding long RNA 付着によるX染色体の不活性化
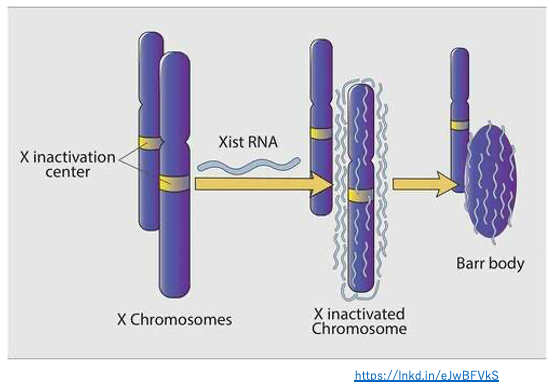
その後の研究で,Xist の転写物であるnon-coding long RNAは,それを転写しているX染色体のほぼ全域に付着(擬似(偽)常染色体領域などを除く)していることが示されました。
15. XIC内にコードされている2つのnon codong long RNA : Xist とTsix
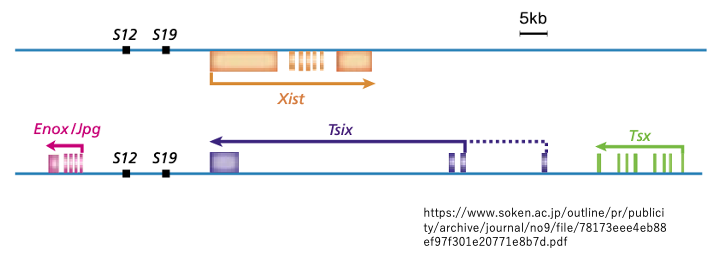
その後の研究で,XIC領域にはXist non coding long RNA遺伝子の他に,その相補鎖からのアンチセンス転写物として,Tsix non coding long RNAを転写する遺伝子が存在していることがわかりました。
また,Tsix RNAはXist RNAの発現を抑制する働きを持つことが分かりました。
16. XIC領域の対合による不活性化X染色体の決定
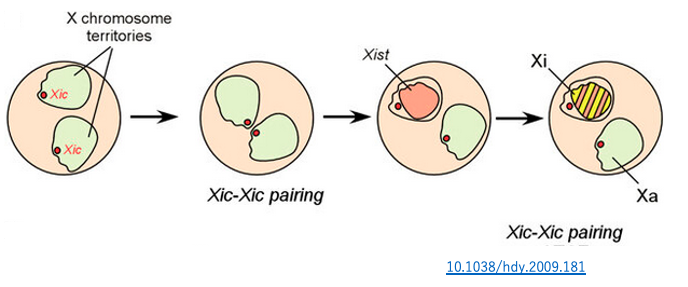
未分化な体細胞においては,両方のX染色体のXistもTsixも低レベルで発現されています。やがて, X染色体が対合するとう現象が確認されています。この時,XIC領域も40分程度対合するのが観察されています。このような一過的なXIC領域の対合現象を通して,ランダムに一方のX染色体の不活性化が決定することが分かってきました。
対合直後に,片方のX染色体でTsix RNAの発現が一過性に抑制されます。抑制的に作用するRNAが低下したことにより,このX染色体ではXist RNA の発現が活性化されます。転写されたXist RNAがそのX染色体に付着することで,X染色体の不活性化が誘導されます。
一方,Tsix RNAの発現量が低下しなかったX染色体では,Xist RNA の発現は低レベルのままなので,不活性化が起きません。
17. XIC付着タンパクの不均衡な配分による不活性化X染色体の決定
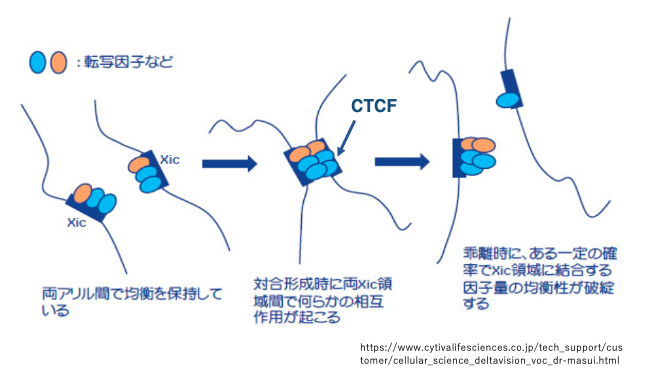
XICは一過的に対合後,分離します。この対合と分離によりXIC領域に付着していた,クロマチン構造の調節に関与するタンパクや転写因子などが不均衡に分配されると,考えられています。
クロマチン構造の調節に関与するタンパクとしては,CTCF (CCCTC-binding factor)が注目されています。
18. ヒストン修飾とDNAのメチル化を伴うX染色体の不活性化
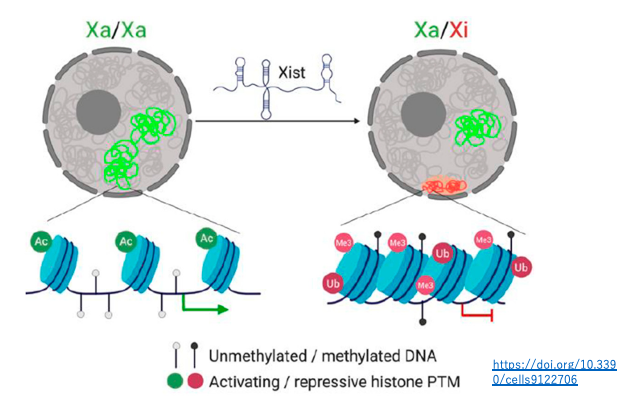
不活性化されたX染色体では,Xist RNAが表面を覆っている他,転写抑制的なヒストン修飾(H3K27me3、H2AK119ub)の頻度が高い,ヒストンのアセチル化頻度が低い,DNAのメチル化率が高いなどの特徴があります。
このような,エピジェネティックなクロモソームの修飾により,不活性化されたX染色体の凝集が安定的に維持されます。
19. 不活性化X染色体が決定される過程
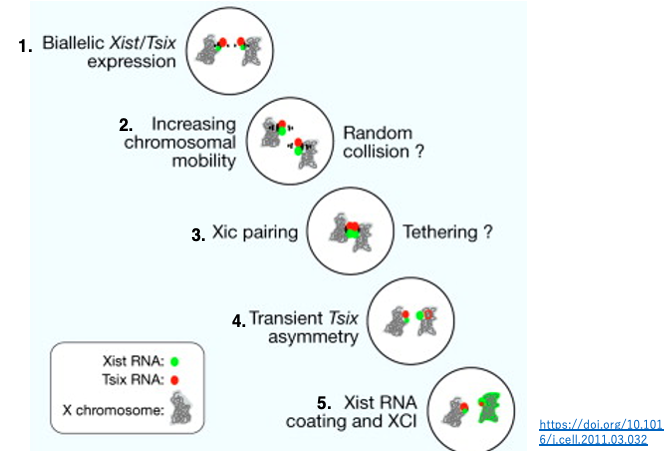
1. 未分化な細胞においては,両X染色体のXIC領域にあるXistとTsix遺伝子では,低レベルの発現が起こっている。X 染色体は凝集していない。
2. X染色体の細胞内での動き(流動性)が活発化して,X染色体とX染色体の衝突が促される。
3. 2つのX染色体のXIC部分のペアリングが短時間起きる。
4. X染色体が分離し,CTCF や転写因子の不均衡な分配が誘導される。その結果,一方のX染色体でTsix RNAの発現が抑制される結果,Xist RNAの発現が活性化し,多量のXist RNAが転写される。
5. Xist RNAを多量に発現しているX染色体では,Xist RNAによって表面全体が覆われ,染色体の高度な凝集が始まりまる。ただし,擬似(偽)常染色体領域などでは,Xist RNAの付着ははく,転写が可能な状態となっている。
コメント