この講義の内容は下記のYouTubeサイト(再生リスト:Life Science Lectures for You ・動画による生命科学講義)で動画として視聴できます
https://youtu.be/g4TVaSaxkKA
1. 講義内容の概要
多くの動物では,メスがオスから精子を受け取ることなく,正常なメスの個体を発生させることができます。これを単為発生と言います。
しかし,哺乳類ではメスとオスの常染色体にある遺伝子の一部は発現が抑制されています (ゲノムインプリンティング)。そのため,発現可能な遺伝子をone set揃えるためには両性由来のゲノムが必須となっています。
Key words: 単為生殖, 3倍体銀ブナ, ミツバチの生活環, Paternally Expressed Gene (PEG), Maternally Expressed Gene (MEG), Conflict Hypothesis (対立仮説)
2. 単為生殖 (Parthenogenesis)

メスが精子と受精をせずに,新しいメス個体を産む生殖方法は単為生殖 (Parthenogenesis)と呼ばれます。
メリットとしては,交尾相手を見つける必要がないので,繁殖にかかるエネルギーと時間を節約することができ,迅速に個体を増やすことができる。
ディメリットとしては,親と子,子の間での遺伝的多様性が低くなり環境が変化した時の適応力が低下した集団に成りやすいと考えられています。
3. 3倍体銀ブナの単為生殖
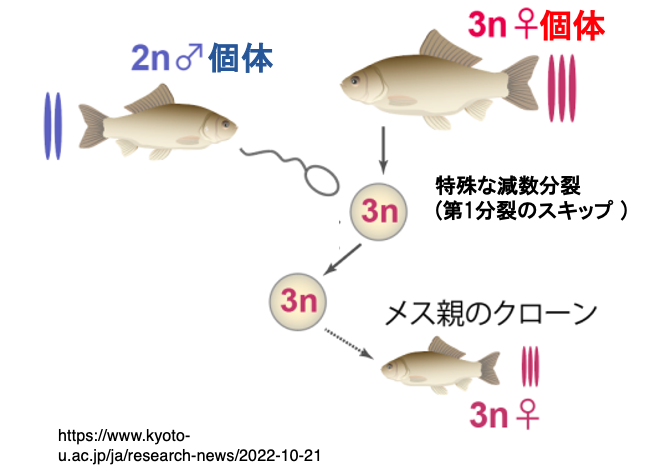
銀ブナのほとんどの個体は3倍体のメスです。3nの卵原細胞は特殊な減数分裂(第1分裂のスキップ)を経て3nの卵が生成されます。
卵は近縁種の2nのフナなどの魚の精子によって賦活化されて発生を開始しますが,精子の核は排除されます。そのためメス親と同じ遺伝子構成を持つメスの子が作られることになります。
したがって,3倍体銀ブナは単為発生により個体を増やしており,個体間の関係性はクローンということになります。
4. 単為生殖を含むミツバチの生活環
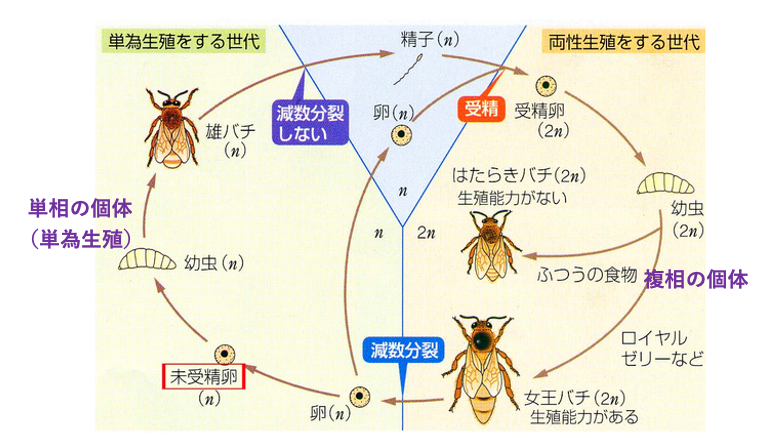
ミツバチでは2nの女王バチのみが生殖能力があり1nの卵を産みます。この卵が精子と受精することなく発生すると1nのオスとなります。オスは精子形成過程で減数分裂を行わずに1nの精子を形成することができます。ミツバチのオスは単為生殖で生まれます。
ミツバチでは2nの個体は,すべてメスとなります。メスの幼虫がロイヤルゼリーを与えられずに発生すると,2n の働きバチとなりますが,生殖能力がありません。ロイヤルゼリーを与えられた1匹のメスのみが生殖能力を持つ女王バチになり,卵を生産することができます。
女王バチはオスと交尾することで,2n の受精卵を形成できます。2nの受精卵は,発生すると全てメスになります。ミツバチでは1倍体の個体はオス,2倍体はメスとなります。
5. 卵と極体の融合による単為生殖:Automixis
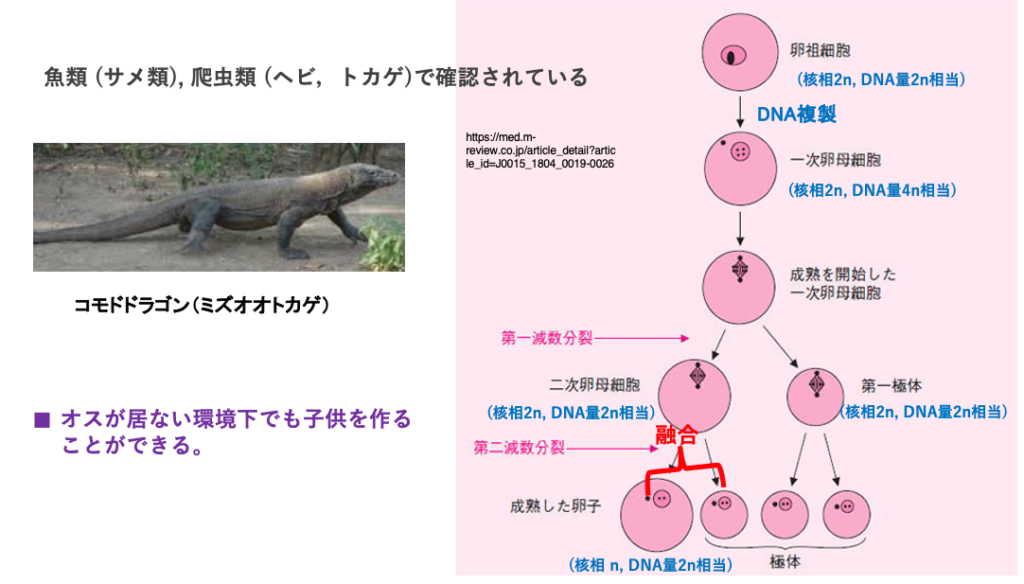
単為生殖には,3倍体銀ブナのようにクローンを作って増殖する他に,魚類や爬虫類では卵と極体の融合により,2n の卵が形成され,それが発生して個体が形成されるタイプの単為生殖,Automixisがあります。
この場合,卵母細胞の中にあった相同染色体の組み合わせが変化するので,メス親とメスの子では,染色体構成が同じではないので,クローンではありません。また子供達の間でもある程度の遺伝的多様性が確保されます。
このような単為生殖では,オスが居ない環境下でも子供を作ることができるというメリットがある一方で,生まれる子供の遺伝的多様性が限定的であるとうディメリットがあります。
6. 単為生殖がブロックされている生物
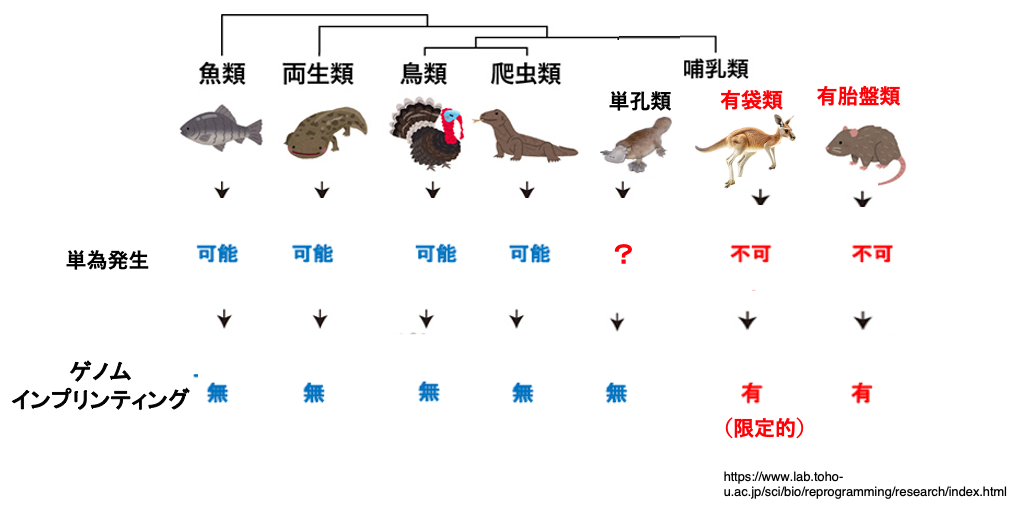
全ての生物で単為生殖が可能ではなく,有袋類のカンガルーや発達した胎盤を持つ有胎盤類では,遺伝的にそれをブロックするシステムが有ります。
有袋類や有胎盤類では,メスとオスの常染色体にある遺伝子の一部は予め発現が抑制されています。これをゲノムインプリンティングと言います。そのため,発現可能な遺伝子をone set揃えるためには両性由来のゲノムが必須となっています。
7. 単為生殖をブロックするゲノムインプリンティング
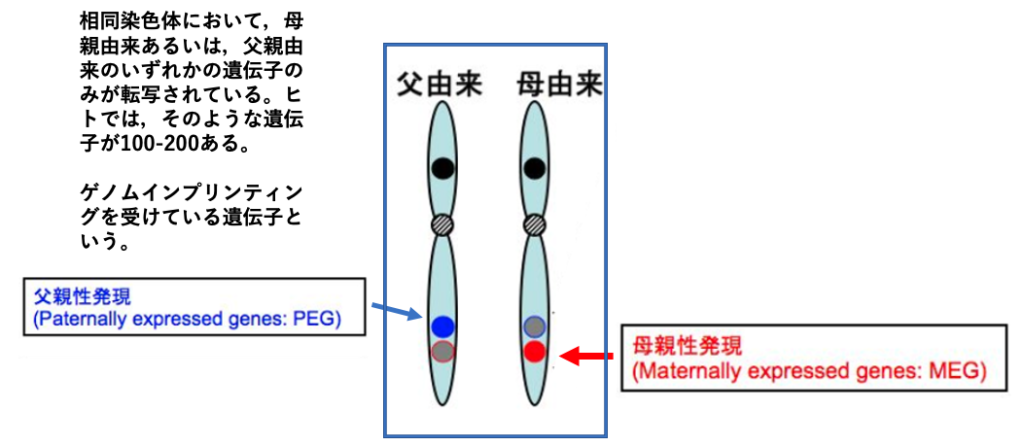
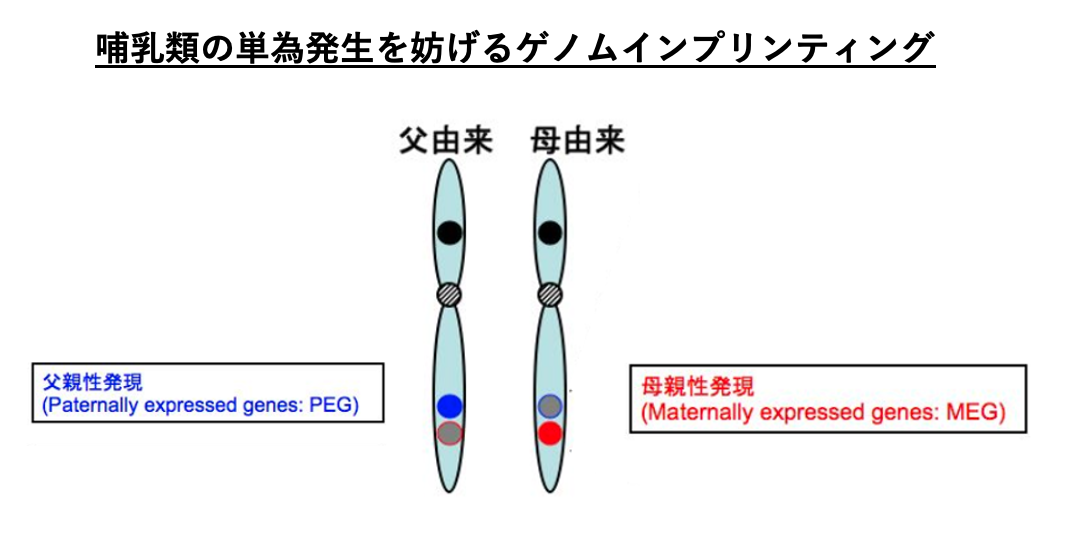
相同染色体において,母親あるいは父親由来のいずれかの対立遺伝子のみが転写されている現象は,ゲノムインプリンティングと呼ばれます。
相同染色体において,父親由来のゲノム上の遺伝子は転写されていて,母親由来の対立遺伝子が転写されていない場合,転写されている父親由来のゲノムにある遺伝子はPaternally Expressed Gene (PEG)とよばれ,その逆で,母親由来のゲノムでのみ発現されている対立遺伝子はMaternally Expressed Gene (MEG)と呼ばれます。
8. DNAのメチル化によるゲノムインプリンティング
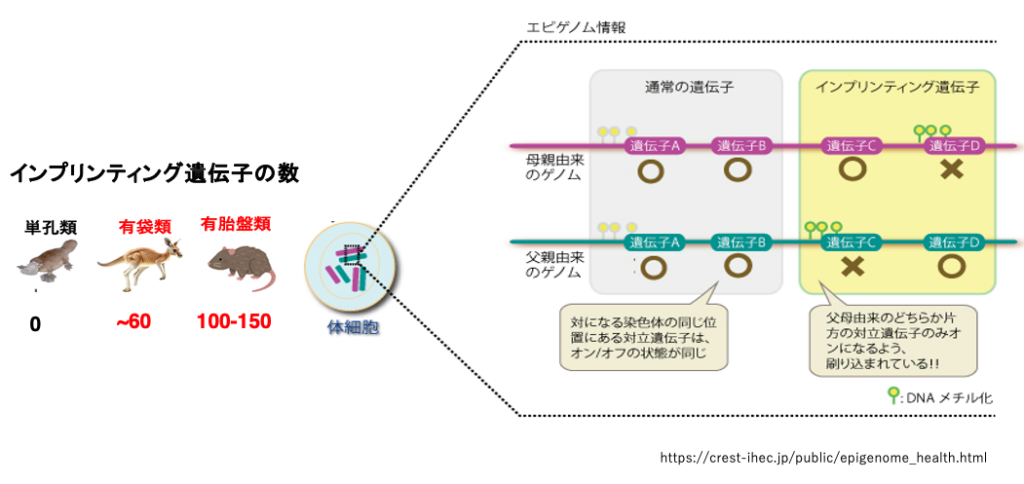
未発達な胎盤を持つ有袋類では,このようなゲノムインプリンティングを受ける遺伝子の数は60個程度と推定されています。発達した有胎盤類では100〜150個の遺伝子がゲノムインプリンティングを受けていると推定されています。このような,特定の遺伝子の発現抑制はDNAのメチル化とヒストン修飾により,体細胞では終生維持されます。一方,胎盤を持たず,卵を産む単孔類では,そのようなゲノムインプリンティングは無いと推定されています。
したがって,有袋類や有胎盤類では爬虫類の例のように,卵と極体が融合して2nの卵ができたとしても,両方のゲノムが母親由来であるため,Paternally Expressed Gene (PEG)の発現が欠落する結果となり,胚の発生は妊娠のごく初期で停止してしまいます。
9. PEG・MEGの染色体間分布
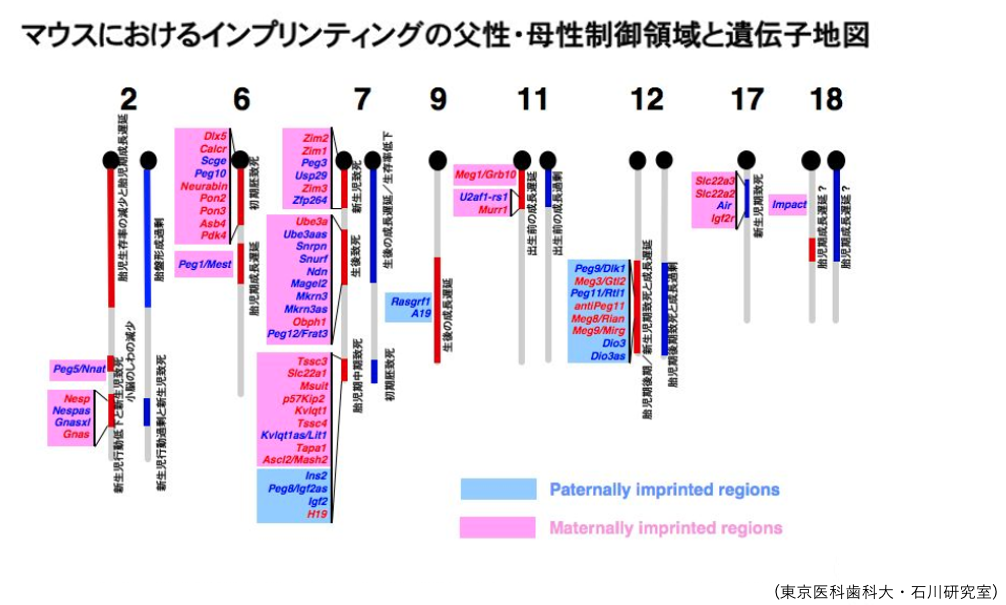
Paternally Expressed Gene・Maternally Expressed Geneは特定の染色体の一部に集中してクラスターを形成しています。
これにより,エピジェネティックな制御機構(DNAメチル化やヒストン修飾など)が効率的に適用されることになります。
10. 刷り込み直し
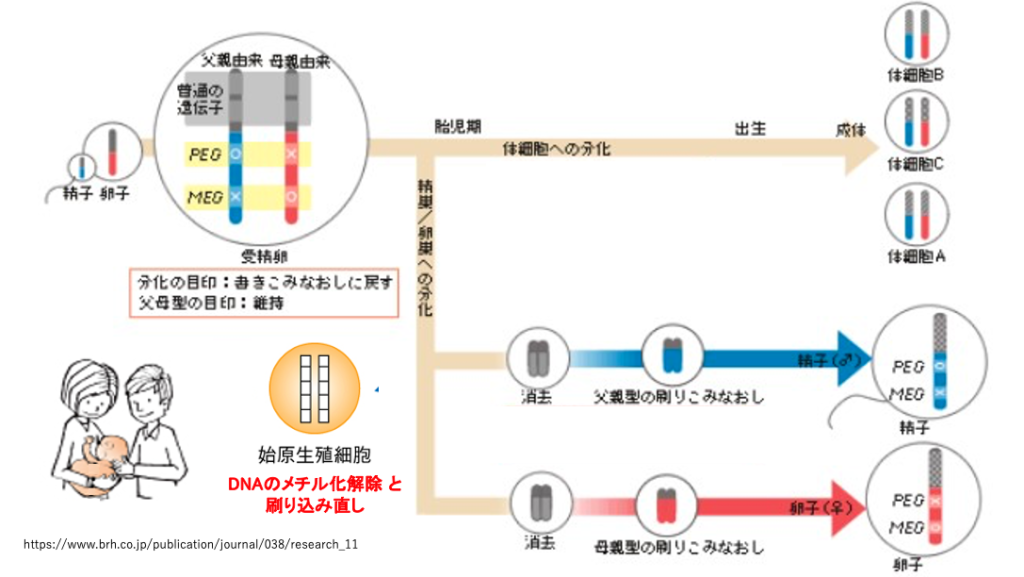
発現抑制はDNAのシトシンのメチル化とヒストン修飾というエピゲネティックな機構によります。そのため,体細胞において発現抑制は終生継続されます。
一方,次世代の個体が生産する精子や卵では,PEG/MEGがリセットされて,それぞれの性に適合してものに刷り込みなおされる必要があります。
胎児の始原生殖細胞内においては,PEG/MEG の解除に必要なDNAのメチル化消去が性別によって異なる時期に起きます。また,それぞれの性に適合した刷り込みは,オスでは受精後15-16日目以降に,メスでは出生後2―3日目以降に起こります。
11. ゲノムインプリンティングの役割

現状,ゲノムインプリンティングの役割については決定的な説がありません。初期に出された仮説としては,対立仮説があります。
これは,“胎生という生殖様式を示す動物では、雄は自分の子供を大きくするように、一方、雌は1回の出産の負担を減らし機会を増やすために子供を小さくするように進化上の圧力がかかる。つまり、父親性発現遺伝子は胎児の成長促進を、母親性発現遺伝子は、成長抑制に働く”という仮説です。しかし,研究の進展に伴い明らかになったインプリント遺伝子が増えるにつれ、コンフリクト仮説の例外が増えています。
PEGの代表的な役割としては,胎盤形成や正常な胎盤機能の維持に関与が知られています。MEG の代表的な役割としては,胎児の成長を抑制する役割を持つものや,胎児や新生児の代謝プロセスの調節,神経系の発達,脳や神経系の正常な形成に関与するものがあります。
コメント