この講義の内容は下記のYouTubeサイト(生命科学を専攻する学生の為の分子生物学講義)から視聴可能です
https://www.youtube.com/watch?v=3WOlTxRv_m4&list=PL_B52Q_vHW1Y0YgqMBs_HZXBbtISm3L7B
1. 講義内容
このサイトでは以下の項目について解説をしています。29枚の図が含まれています。
- tRNA分子の特徴と役割:CCAの2’-OHまたは3’-OH末端へのアミノ酸付加
2. アミノアシル化tRNA生成の為の2つのStep: ARS酵素によるAminoacyl AMP生成とAminoacyl tRNA生成
3. tRNAに受容されたアミノ酸の化学修飾による変換
4. 遺伝暗号表の特徴:Two codon set, Family box
5. Wobble base pair: アンチコドン1番目とコドンの3番目の水素結合形成の特別ルール
6. 終始コドンはどのようにして翻訳を終結させるのか:Release factor (翻訳終結因子)による終始コドンの認識
7. 翻訳の開始に関与するInitiator tRNAと受容アミノ酸:fMetを受容するInitiator tRNAの生成
Key Words:
2. tRNA: transfer RNA (運搬RNA)
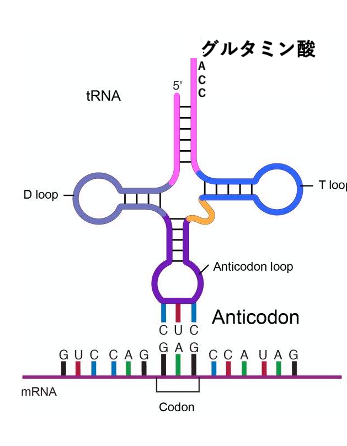
まず初めに,tRNAにアミノ酸が付加されるメカニズムと遺伝暗号表の特徴について説明をします。
tRNAは特定のアミノ酸をメッセンジャーRNAの特定のコドンまで運ぶ、 80塩基程度の一本酸RNAです。 その3’末端は必ずCCA配列で終わるという特徴を持っています。 また、アンチコドンループ領域内には,メッセンジャーRNA上のコドンと対合して 安定な水素結合を形成できるアンチコドンがあります。 この図では、アンチコドンCUCを持つtRNAが,グルタミン酸を3’末端に結合し,メッセンジャーRNA上のコドンGAGと安定な水素結合を作っている様子を示しています。
3. Aminoacyl-tRNA synthetase : 特定のアミノ酸を特定のトランスファーRNAに結合させる酵素
特定のアミノ酸を特定のトランスファーRNAに 使用させるような活性を持つ酵素がAminoacyl-tRNA synthetaseです。略して,ARSと呼ばれます。 細胞内で使われるアミノ酸の数は20種ありますので、ARSは原則として20種あります。
それに対してtRNAは、内部の塩基配列が違う場合を,異なる種類のトランスファーRNAだという風に数えた場合、40から60種類あることが分かっています。
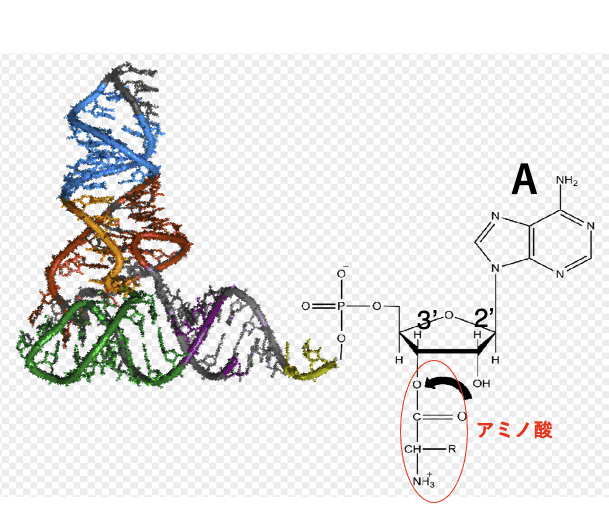
ARSは,アミノ酸を付加させるリボースの位置の違いにより,2つに大別されます。 Class I-ARSと呼ばれるものは、tRNAの3’末端配列であるCCAのアデノシンのリボースの2’-OHにアミノ酸を付加させる活性を持ちます。 それに対してClass-II ARSは、アデノシンのリボースの3’-OHにアミノ酸を付加させる活性を持ちます。 このように、アミノ酸はtRNAの末端のアデノシンの2’-OHあるいは3’-OHに付加されます
4. 2種類のAminoacyl-tRNA synthetase (ARS)
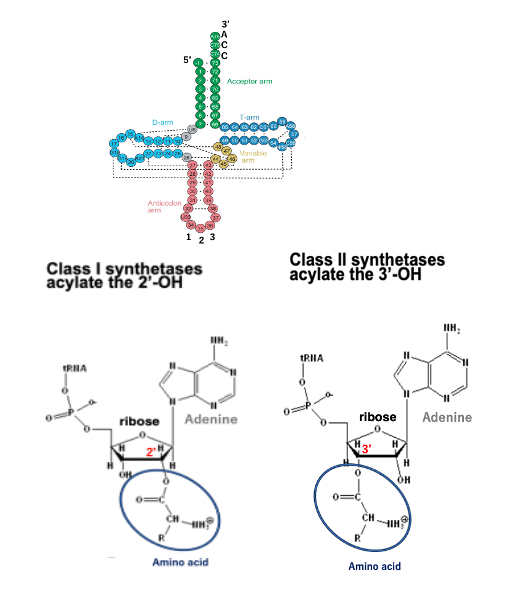
特定のアミノ酸を特定のトランスファーRNAに 使用させるような活性を持つ酵素がAminoacyl-tRNA synthetaseです。略して,ARSと呼ばれます。 細胞内で使われるアミノ酸の数は20種ありますので、ARSは原則として20種あります。 それに対してtRNAは、内部の塩基配列が違う場合を,異なる種類のトランスファーRNAだという風に数えた場合、40から60種類あることが分かっています。 ARSは,アミノ酸を付加させるリボースの位置の違いにより,2つに大別されます。 Class I-ARSと呼ばれるものは、tRNAの3’末端配列であるCCAのアデノシンのリボースの2’-OHにアミノ酸を付加させる活性を持ちます。 それに対してClass-II ARSは、アデノシンのリボースの3’-OHにアミノ酸を付加させる活性を持ちます。 このように、アミノ酸はtRNAの末端のアデノシンの2’-OHあるいは3’-OHに付加されます。
5. 各Class I, Class II ARSが担当するtRNAのまとめ
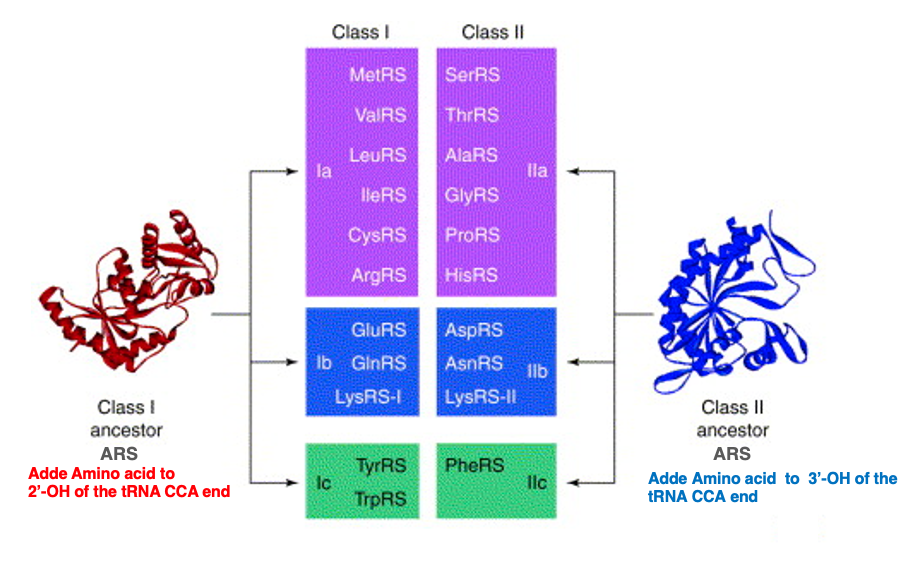
この表はClass IのARSとClass IIのARSをまとめたものです。 メチオニントランスファーRNAにメチオニンを孵化することができるようなARS、 それからバリン、ロイシン、イソロイシン、システン、アルジニン、 グルタミン酸、グルタミン酸,リジン,tyrosine、トリプトファンを tRNAに付加するようなARSはClass I、 つまり,riboseの2’-OHにアミノ酸を付加するという活性を持っています。
一方、こちら側にあるのはClass II ARSで、riboseの3’-OH末端にアミノ酸を付加します。ARSが特定のトランスファーRNAを認識して、 特定のアミノ酸を付加する反応は2ステップの反応で起こるということが知られています。次に,この反応について説明をします。
6. アミノアシルAMPが形成
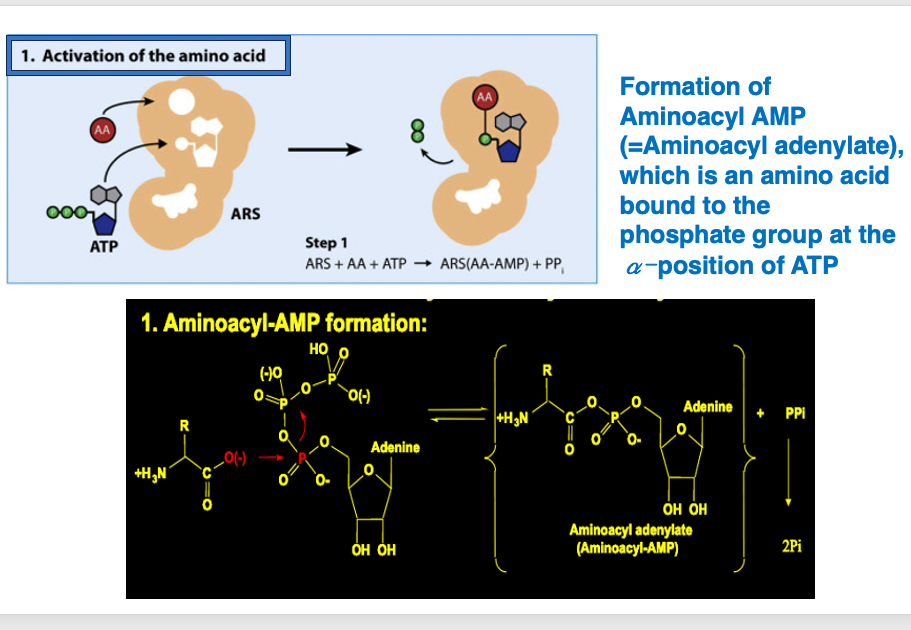
まず一番目の反応から説明をしていきます。 一番目の反応として ARSは ATPと自分自身が 取り込むべきアミノ酸を認識して, それをARS内に取り込みます。 ARSの中で ATPのα位のリン酸基に アミノ酸が付加された アミノアシルAMPが形成されます。
化学式で書くとこうなります。 ATPのα位のリン酸基に アミノ酸が付加されて アミノアシルAMPが形成され,ピロリン酸が放出されます。
7. tRNAへのaminoacyl AMP転移
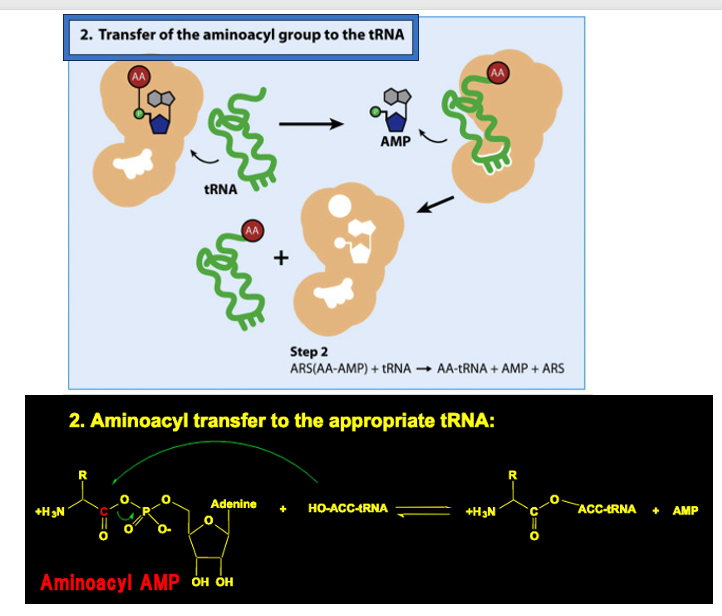
2ステップ目の反応として,ARSは 自分が取り込むべき トランスファーRNAを認識して 酵素内部に取り込みます。ARSはtRNAを内部に取り込むと その2‘-OH あるいは3’-OHに、アミノアシルAMPからアミノ酸を転移させます。
この反応を化学式で書くと,このように表現されます。
Aminoacyl AMPが持っていたアミノ酸がtRNAに渡されると同時にAMPが放出されています
8. tRNAへのアミノ酸付加反応のまとめ
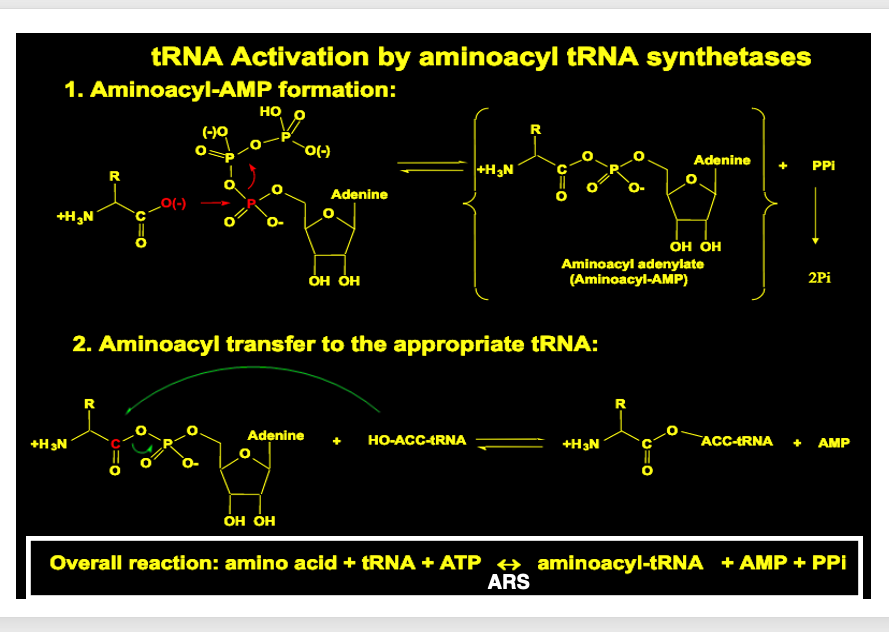
Step 1とStep 2, 2つの反応を化学式でまとめますと,このようになります。アミノ酸とtRNA, ATPが反応に関与します。ARSは,その反応を触媒します。
最終的に 特定のtRNAに 特定のアミノ酸が付加された tRNA分子が 形成され,AMPとピロリン酸が 排出されます。
9. tRNAの種類を識別する配列 (tRNA Identity)
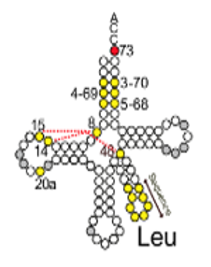
Leuを受容するtRNAは複数種類あり、その内部塩基配列は異なります。しかし、Leu-tRNAに結合させるアミノアシルtRNA合成酵素は1種類しかありません。では、Leu-ARSはどのようにしてLeu-tRNAを他のtRNAと区別しているのでしょうか?
Leu-tRNAの識別について説明しましょう。
10. ARSはどのようにtRNAを識別しているのか
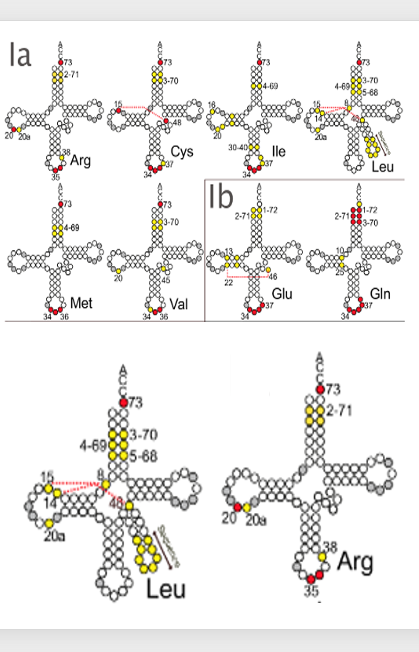
次に、ARSはどのようにtRNAを識別しているのかに関して解説をしていきます。 例えば、ロイシンを運ぶべきトランスファーRNAは細胞の中に複数種あって、その内部の塩基配列は同じではありません。 一方、ロイシンをトランスファーRNAに付加させるARSは1種類しかありません。従って、ロイシンARSはどのようにしてロイシンtRNAをロイシン意外のアミノ酸を受容するtRNAと見分けているのかということが問題になります。
ここにロイシンtRNAの図がありますが、ロイシンを運ぶべきトランスファーRNAでは、 この73番目の塩基が共通でいつも決まった塩基であることがわかっています。 その他、この黄色で塗ってある塩基はロイシンtRNA間で強く保存されています。 このような塩基をtRNAのアイデンティティを決める塩基というふうに言います。
例えば、これはアルギニンtRNAですけれども、アルギニンを運ぶtRNAだと、 この73番目、20番目、この35番目、6番目、このような塩基がアルギニンtRNA間で共通であり,アイデンティティ塩基として使われているということがわかります。 ARSは各tRNAのアイデンティティを決める塩基を強く認識して、 どのアミノ酸を運ぶトランスファーRNAであるかを認識しています。
11. tRNAに受容されたアミノ酸の化学修飾
一部のバクテリアではアスパラギン酸tRNA合成酵素を持っていますが、アスパラギンtRNA合成酵素を欠いています。同様に、グルタミン酸tRNA合成酵素を持っていますが、グルタミンtRNA合成酵素を欠いています。このような状況をどのようにして,解決しているのでしょうか。
解決法補として,tRNAに受容されたアミノ酸の化学修飾が行われていることが分かっています。
12. アスパラギンtRNAへのアスパラギン酸の付加とその後の化学修飾
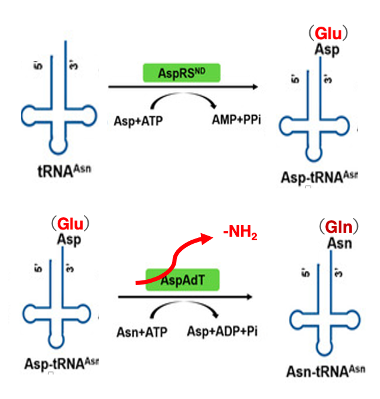
アミノ酸が受容されたtRNAの上で化学修飾を受けて,受容されているアミノ酸が他のアミノ酸に変換されるという例についてお話をしていきます。
一部の細菌等ではグルタミン酸ARSは存在するがグルタミンARSは存在しない, あるいはアスパラギン酸ARSは存在するが,アスパラギンARSが存在しないことが知られています。このような場合,tRNA上でアミノ酸を変換することで,これに対応しています。これはアスパラギンを受容すべきtRNAがアスパラギンARSはこの細菌ではないので 一旦アスパラギンtRNAにアスパラギン酸が付加されます。
従って,アンチコドンから見たtRNAの種類と付加されたアミノ酸の間で不一致があるという状態になっています。 それを解消するために,アスパラギン酸からアミノ基を一つ取り除くことで,アスパラギン酸をアスパラギンに変換します。これによって,アスパラギンを受容したアスパラギtRNAが細胞内に生まれます。
同様に一部の細菌ではグルタミンARSがありません この場合グルタミンtRNAに一旦,グルタミン酸を付加させた後に,おいてグルタミン酸からアミノ基を,一つ取り除くことでグルタミンを受容したグルタミンtRNAが細胞内に出現することができます。
13. Met をfMet, SerをセレノシステインにtRNA上で変換
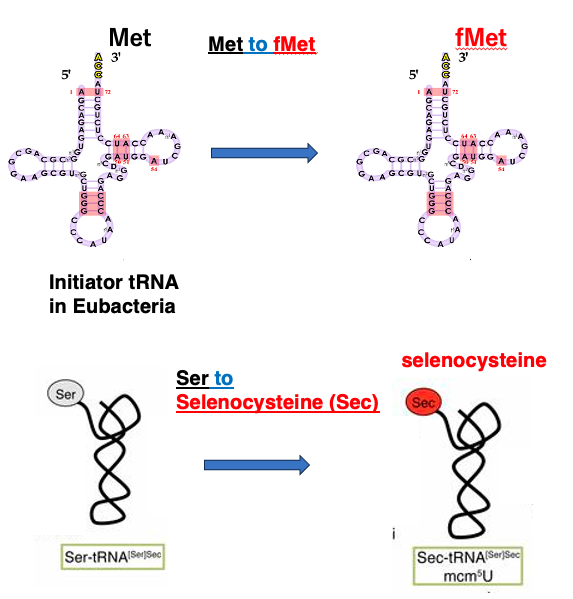
もう少し同様の例についてお話をします。
真正細菌のイニシエーターtRNAでは、 まずメチオニンが受容されて、 その後にメチオニンがtRNA上で、 フォルミルメチオニンに変換されます。
また、セレノシステインtRNAの場合は、 まずセリンがセレノシステインtRNAに付加されます。付加されたセリンがtRNA上で、 セレノシステインに変換されます。
14. 地球上の生物すべてに適用可能な遺伝暗号表
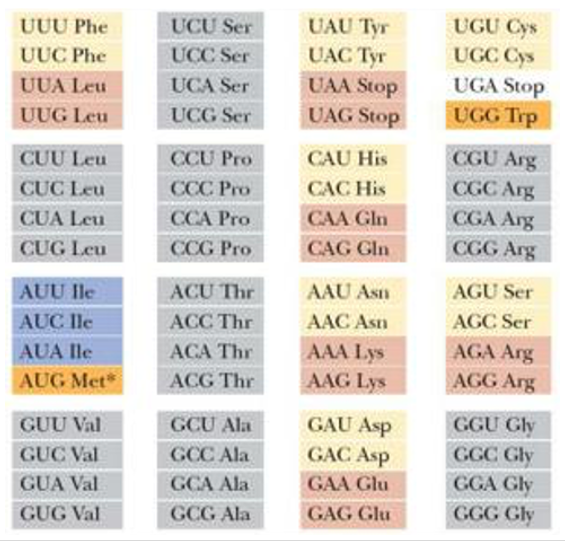
地球上の生物であれば,細菌,動物,植物,ウイルスの如何を問わず,使用するアミノ酸は共通で,しかも20種類に限られています。一方,コドンと呼ばれる連続する3塩基が,一つのアミノ酸に対応するシステムとなっています。連続する3塩基には64種類ありますから,20種のアミノ酸を指定する場合,コドンとアミノ酸の対応関係は1対1とならず,コドンが違っても指定されるアミノ酸が同じという場合が発生します。
また,この遺伝暗号表は,基本的には原核生物にも真核生物にも適用することができます。
基本的にはといったのは,様々な生物種のミトコンドリアについて,コドンとアミノ酸の対応関係が,一部においてこの遺伝暗号表とは違っています。更に, 一部の原核生物や真核生物でも この遺伝暗号表からずれた対応関係がある場合が知られています。しかし,そのズレは,2次的に,進化の途中で生じたものであることが分かっています。
15. 遺伝暗号表の特徴
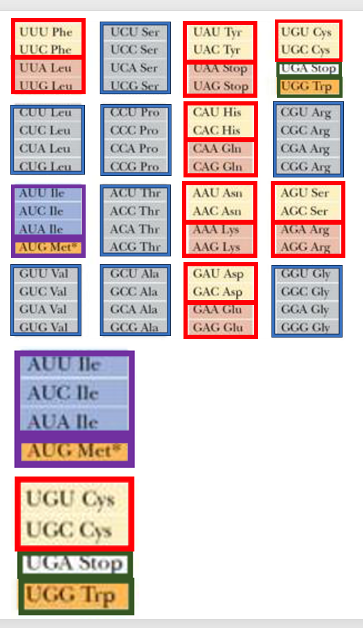
これはでは次に特定のコドンに特定のアミノ酸が受容されたtRNAが対合するためのルール, コドンとアンチコドンの対合ルールについて解説をしたいと思います。 ここに示したのが遺伝暗号表です。 この遺伝暗号表の見方ですけれども, UUUというコドンがあると,それにはアミノ酸としてフェニルアラニンが対応するというふうに解釈します。これは, コドンUUUはフェニルアラニンを受容しているtRNAが そのアンチコドンを使って, コドンUUUと安定な水素結合を形成するというメカニズムのために実現しています。
遺伝暗号表の特徴ですけれども コドン3番目の塩基がUでもCでも対応するアミノ酸が同じ, あるいはAでもGでも対応するアミノ酸が同じという箇所がたくさんあります。このような冗長性のあるコドンに対するアミノ酸の割り当て方式をTwo-codon set言います。 フェニルアラニンもTwo-codon setですし,このロイシンもTwo-codon set, アスパラギン酸、グルタミン酸についてもTwo-codon set方式のアミノ酸assignmentであることが分かります。
その他にコドン3番目がU、C、A、G どれであっても対応するアミノ酸が同じになるような,アミノ酸の割り当て方式があります。これは,family boxと呼ばれます。例えばこのロイシンのコドンがそうですね。 コドンの1番目の塩基がC、2番目がUであれば, 3番目の塩基はU、C、A、G どれであっても,対応するアミノ酸はロイシンとなっています。 コドン表の多くの部分は,Two-codon setあるいはfamily boxで構成されていますが,例外的な部分も存在します。 このイソロイシンのBoxがそうですね。コドンの3番目の塩基が U、C、Aの場合がイソロイシンで Aの場合はメチオニンというふうに3対1に分かれています。 それからこの部分ですね。 システインのassignmentに関しては,Two-codon setとなっていますが, UGAだと終始,UGGだとトリプトファンが対応しています。これら,2箇所については,非常に特殊な分割がされているということが分かります。
16. RNA鎖間で形成される水素結合
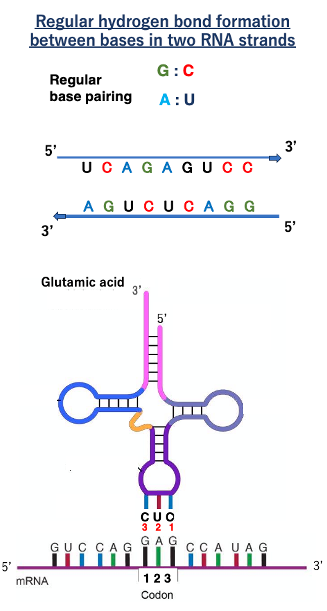
まず,RNA鎖間における通常の水素結合について説明します。逆方向に2本のRNA鎖が向き合うことで,塩基間で水素結合を形成できます。GはCと,AはUとの間で安定な水素結合を形成できます。
これはグルタミン酸を受容したtRNAが コドンGAGと安定な水素結合を作っているという図です。ここにあるのがメッセンジャーRNA。 こちらが5’-端、こちらが3’-端です。 核酸については基本的には,5’-端から3’-端に向かって順番をつけますので ,コドンGAGの1番目の塩基はG、2番目がA、3番目がGということになります。 このコドンと安定な水素結合を作っているtRNAのアンチコドンは CUCです。アンチコドンの1番目がC,2番目がU,3番目がCです。
塩基と塩基が安定な水素結合を作るためには 2つの核酸分子の方向性は逆向きである必要があります 。それはDNA鎖間でもRNA鎖間でも同じです。mRNAがこのような方向を向いているなら tRNAはこのように逆向きになる必要があります。
17. Wobble Rule: 3番目のCodonと1番目のAnticodon間の特殊な水素結合形成
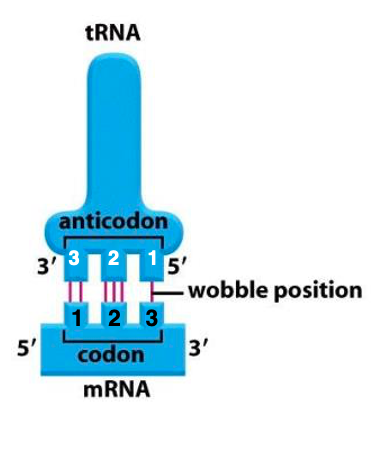
Wobble ruleがどのような点で 特殊かと言いますと,transfer RNAのアンチコドンの 1番目がUの場合 通常のルールであれば Aとしか 結合しないはずが,コドンの3番目が AであってもGであっても対合が許される, ということでます。
また,transfer RNAのアンチコドンが Gの場合は 通常ならCのみですけれども コドンの3番目がUであっても 対合が可能です。 一方,アンチコドンの1番目がCの場合は 通常と同じで コドンの3番目はGのみ, アンチコドンの1番目がAの場合が コドンの3番目のUとしか対合できないというのは,通常と同じです。
18. Anticodon 1番目の塩基修飾と対合:イノシン(I)の場合
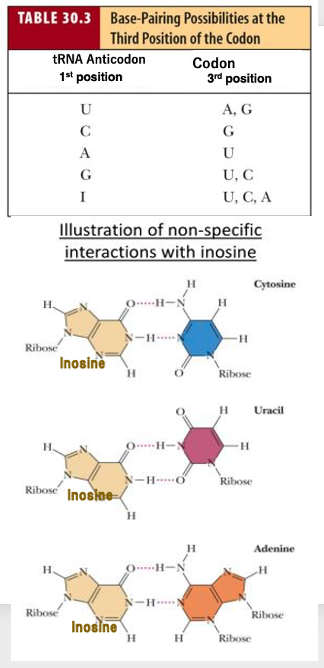
また,transfer RNAの アンチコドンの1番目というのには 様々な 化学修飾を受けた塩基が存在することが知られています。
例えば,アンチコドンの1番目が イノシンですと,コドンの3番目がUであってもCで あってもAであっても 対合ができます。
それがこの例ですね トランスRNAの 1番目の塩基がイノシンの場合 CでもUでもAでも,ここに示したように水素結合が成立するということが分かっています。
19. Two-cond set: Phe Codon, Leu CodonにおけるtRNAとの結合
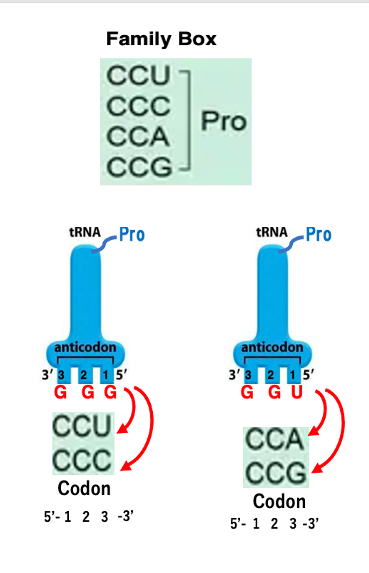
ではもう少し具体的な場合について見ていくことにします。
アンチコドンGAAを持つtransfer RNAにはフェニルアラニンが付加されます。アンチコドンの1番目とコドンの3番目についてはWobble Ruleが成立しますから,このアンチコドンを持つtransfer RNAが 安定な水合を形成できるコドンというのはUUUかUUCというコドンということになります。 したがって,アンチコドンGAAを持つフェニルアラニンtransfer RNAが細胞内に存在する限り,コドンUUUもUUCもフェニルアラニンに対応するということになります。
アンチコドンUAAを持つtransfer RNAにはロイシンが付加されます。 このアンチコドンを持つtransfer RNAが安定な水素結合を形成できるコドンはUUAとUUGということになります。 UUAあるいはUUGコドンにはロイシンtransfer RNAが安定に結合できるので, UUAもUUGもロイシンコドンということになります。
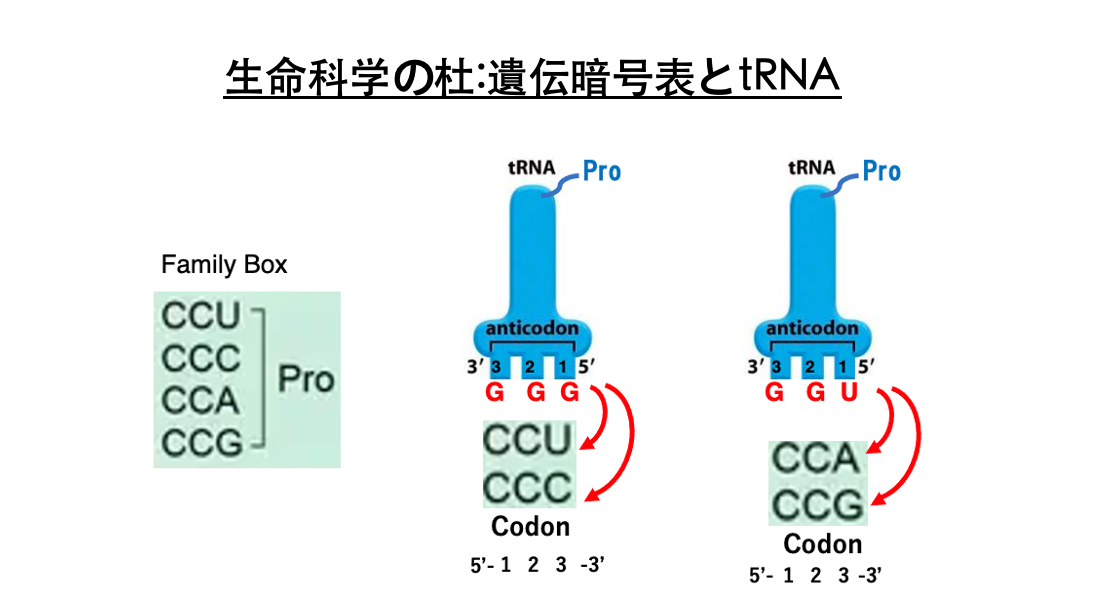
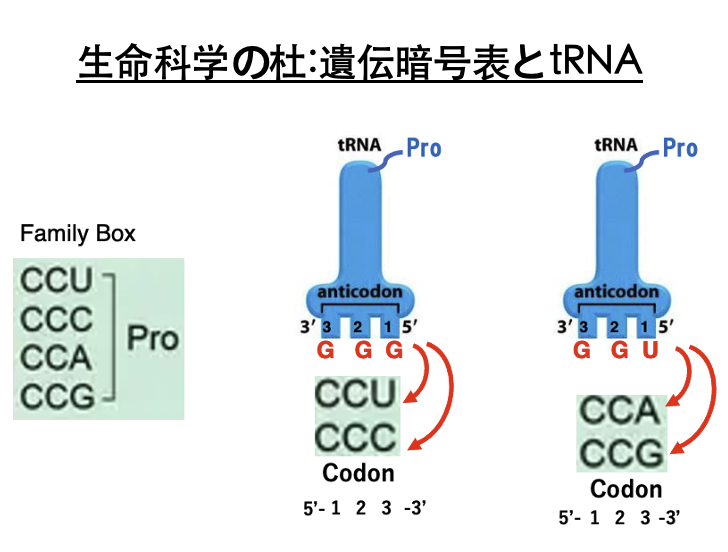
20. Family Box: Pro Codon におけるtRNAとの結合
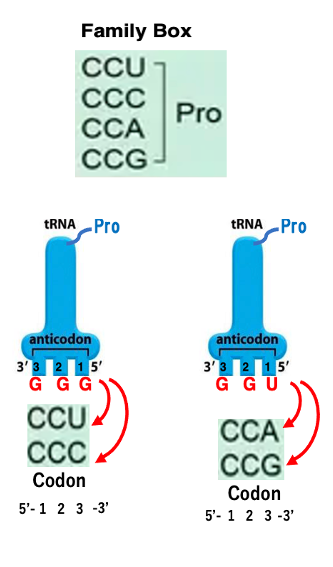
アンチコドンGGGを持つtransfer RNAにはプロリンが付加されます。
このtransfer RNAが安定な水素結合を形成できるコドンはCCUとCCCです。 アンチコドンの1番目とコドンの3番目の間ではWobble Ruleが成立するからでしたね。 またアンチコドンUGGを持つtransfer RNAにもプロリンが同じく付加されます。このアンチコドンを持つtransfer RNAが安定に水素結合できるコドンは,CCAとCCGです。
したがってCCU、CCC、CCA、CCG、3番目のコドンが何であっても,上記の2種類のうちいずれかのプロリンtransfer RNAが対応するということになります。 というので,コドンの1番目がC,2番目がCの場合,コドンの3番目の塩基に関わらず,プロリンを指定することになります。
従って,コドンCCNはプロリンを指定するFamily Boxとなります。
21. Met Codon とTrp Codon: 対応するtRNAのアンチコドンについて
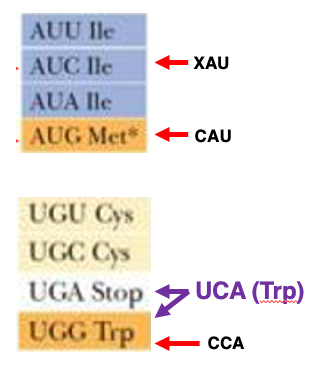
ここはイソロイシンを指定するboxですね。 AUU, AUC, AUA コドンはイソロイシンのコドンです。この3つのコドンのどれとでも安定な水素結合を形成できるtransfer RNAが 存在することが分かっています。そのイソロイシンtransfer RNA Aの アンチコドンの1番目は 非常に特殊な塩基修飾を受けているということが 分かっています。
また AUG メチオニンコドンに 対応するtransfer RNAの アンチコドンの 1番目はCですね。アンチコドンが CAUですと, AUGコドンだけに 対応できるということになります。
また UGG トリプトファン このコドンに 対応する transfer RNAのアンチコドンは CCAということになります 。アンチコドンがCCAだと コドンUGAに対応することはできないので,UGGコドンのみがトリプトファンに対応するということになります。
もしアンチコドン UCAを持つようなトリプトファン transfer RNAが細胞内に出現すると,アンチコドンの1文字目が Uならば AとG の両方と対合することができますので,UGGだけではなくUGAコドンも トリプトファンのコドンとなってしまいます。
22. 終始コドンが翻訳終結シグナルとして機能する理由
UAA, UAG, と UGAコドンは翻訳終結のシグナルとして機能します。これらの3つのコドンに安定的な水素結合をつくれるアンチコドンを持っているtRNAは存在しません。一方,これらのコドンと安定的に結合できるタンパク質が存在します。それらは,Release Factorと呼ばれます。この因子のために,翻訳が終結します。
23. 翻訳のステップ : tRNA上でのペプチド鎖の伸長反応
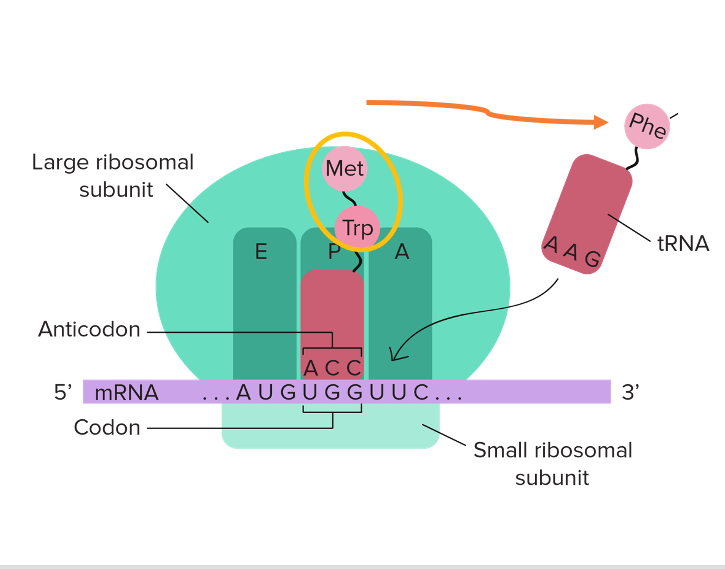
まず、一般的な翻訳のステップについて簡単に説明をしておきます。
リボソームにはtransfer RNAを収容できるような空間が3つあります。 順にEサイト、Pサイト、Aサイトというふうに名前が付けられています。 真ん中のPサイトに入っているtransfer RNAには, 伸びつつある新生ペプチトが付加されています。 その横のAサイトにはアミノ酸を受容したさtransfer RNAが新たに入ってきます。
Aサイトにあるコドンに対応する正しいアンチコドンを持ったtransfer RNAが挿入されますと、 Pサイトにあったtransfer RNAは自分自身が持っていた新生ペプチドをこのAサイトに入ってきたtransfer RNAに渡します。 この反応によって, 新生ペプチドのアミノ酸残基の数が,1つ伸びるわけですね。
この図でいうと、メチオニンとトリプトファンが繋がった2ペプチト鎖に、フェニルアラニンが付加することによって、 3ペプチトにまでアミノ酸残基数が伸びるということになります。
24. 翻訳の終結 : release factorによるペプチド鎖のtRNAからの解離
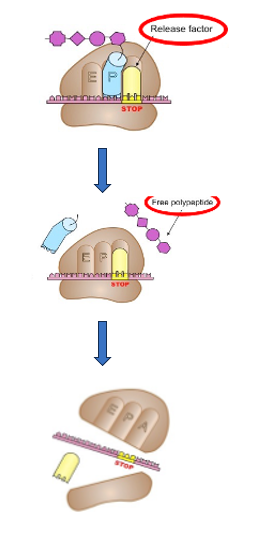
一方、翻訳が終結する場合についてですけれども、今,翻訳が進んできて、ribosomeはそのAサイトに終始コドンが入るというところまで来ています。
Stop codonに対応するようなtRNAは細胞内にはありませんが、 その代わり,release factorがStop codonを認識して、ribosome内で終始コドンと安定な結合を形成します。ペプチジルtRNAのリボースとアミノ酸間のエステル結合が加水分解されることによって、新生ペプチトはtRNAから解離します。新生ペプチトがtRNAのアデノシンを構成するリボースの3’あるいは2’に付加しているのですけれども、そのエステル結合が加水分解を受けることによって、新生ペプチトがtRNAから離されていきます。
それと同時に,GTPのエネルギーを利用してリボソウム、mRNA、release factorの集合体が解体されます。 これが翻訳の終結ということになります。
25. tRNAを擬態するタンパク質 release factor
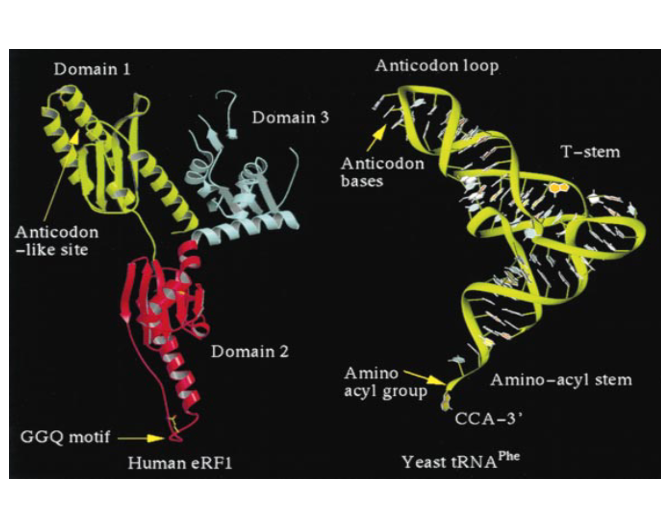
今の説明にでてきた,release factorタンパク分子ですけれども、このタンパクの立体構造(左図)はRNA分子(右図)であるtRNAによく似ているということが知られています。
これは一般的なtRNAの3次元モデルですけれども、このタンパクでできているリリースファクターの立体構造を見てみますと、 その立体構造を作っている分子的な要素は違いますが、2つの分子の立体構造はよく似ているということがわかります。
26. initiator tRNAが受容するアミノ酸
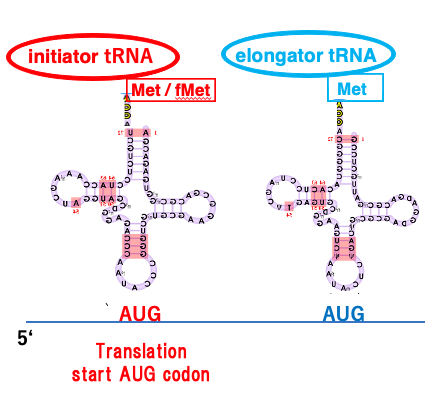
翻訳開始に関与するイニシエーターtRNAが受容するアミノ酸について解説をしていきます。
翻訳開始のコドンはいつもAUGというふうに決まっていますけれども この翻訳開始のAUGコドンに対応するtRNAのことを イニシエーターtRNAというふうに呼びます。
27. initiator tRNAが受容するアミノ酸:真核生物と古細菌の場合
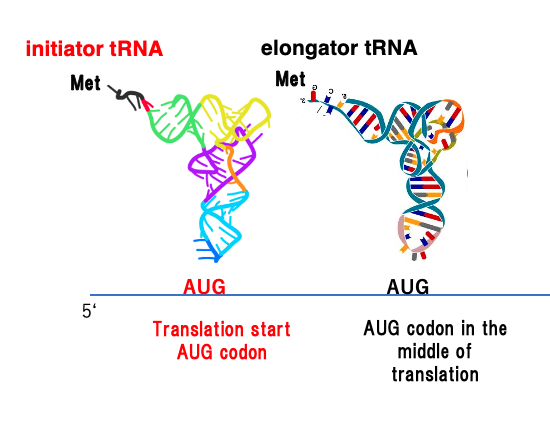
この図は,真核生物や古細菌の場合の翻訳開始を模式化したものです。
翻訳開始の一番初めのAUGコドンにイニシエーターtRNAが対応しますが,イニシエーターtRNAが受容しているアミノ酸はメチオニンです。 翻訳開始ではないAUGコドンがmRNAの途中に出てくることがありますけれども, このAUGコドンを翻訳するのに使われるtRNAは elongator tRNAと呼ばれ, そのtRNAもメチオニンが付加されています。
ということで真核生物や古細菌の場合は ペプチトの先頭のアミノ酸はメチオニンであり, また途中のAUGに対応するアミノ酸もメチオニンということになります。
28. initiator tRNAが受容するアミノ酸:真正細菌,葉緑体,ミトコンドリアの場合
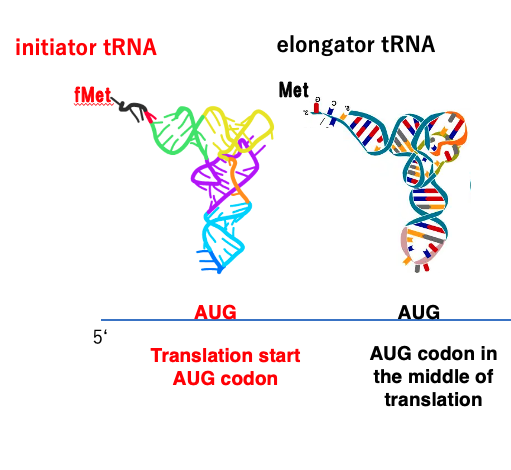
一方、真正細菌や葉緑体ミトコンドリアの翻訳開始は 少し様子が異なります。
これらの生物群においても翻訳開始コドンはAUGですが このイニシエーターtRNAにはメチオニンではなくてフォルミルメチオニンが付加されています。
一方、メッセンジャーRNAの翻訳途中のAUGコドンに対応するのはelongator tRNAですが このtRNAにはメチオニンが付加されています。
29. initiator tRNA上でのアミノ酸修飾:Met をf-Metに変換
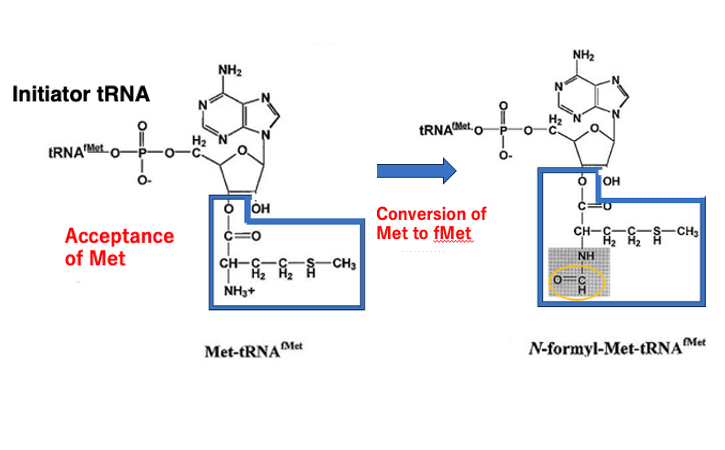
この真正細菌におけるイニシエーターtransfer RNAのformylmethionine受容について解説します。
このイニシエーターtransfer RNAにまずメチオニンが付加されることが分かっています。 その後、transfer RNA上で, メチオニンはフォルミル化を受けてフォルミルメチオニンに変換されるということが分かっています。
一部の真正細菌ではグルタミン受容transfer RNA, アスパラギン受容transfer RNAは,まずはアスパラギン酸、あるいはグルタミン酸を受容して,そのアミノ酸が transfer RNA上で,グルタミンやアスパラギンに変換されるという話をしましたが, このイニシエーターtransfer RNAのフォルミルメチオニン受容に対しても,transfer RNA分子上でのアミノ酸修飾が起こっていることになります。
では,これでこの講義を終わります。
コメント